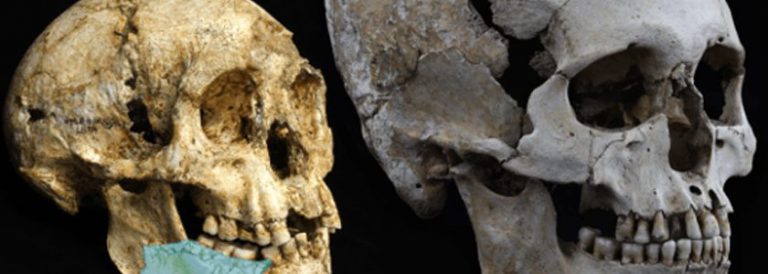
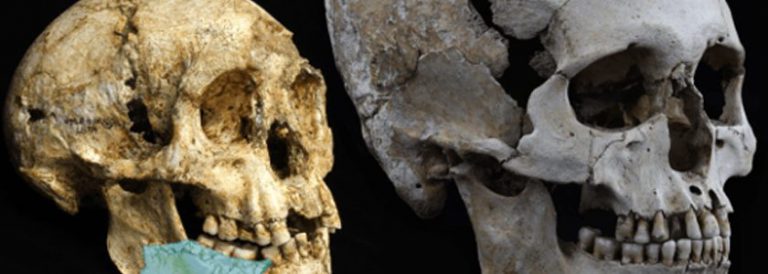
16. Els hominins: els australopitecins
24 novembre, 2014Els primers homínids dels quals es té la seguretat que van ser completament bípedes són els membres del gènere Australopithecus, dels quals s’han conservat esquelets molt complets (com el de la famosa Lucy). Avui parlarem dels australopitecins.
Aquest tipus d’hominí va prosperar a les sabanes poblades d’arbres de l’est d’Àfrica entre 4 i 2,5 milions d’anys enrere amb notable èxit ecològic, com ho demostra la radiació que va experimentar, amb almenys sis espècies diferents escampades des d’Etiòpia i el Txad fins a Sud-àfrica: Australopithecus anamensis, Australopithecus afarensis, Australopithecus africanus, Australopithecus garhi, Australopithecus bahrelgazhali i Australopithecus sediba.
La seva desaparició s’ha atribuït a la crisi climàtica que es va iniciar fa uns 2,8 milions d’anys i que va conduir a una desertificació de la sabana, amb la consegüent expansió dels ecosistemes oberts, esteparis. Com a resultat d’aquesta pressió evolutiva alguns Australopithecus es van especialitzar en l’explotació de productes vegetals durs i d’escàs valor nutritiu, desenvolupant un impressionant aparell masticatori, originant el gènere Paranthropus; altres Australopithecus es van fer gradualment més carnívors, originant els primers Homo.
Les característiques principals del gènere Australopithecus són:
Al crani:
- Capacitat cranial relativament petita.
- Parets cranials primes.
- Vores supraorbitàries robustes.
- Protuberància occipital externa per sota del pla de Frankfurt.
- Flexió basicranial.
- Posició anterior dels còndils occipitals.
- Orientació del foramen magnum en un pla gairebé horitzontal.
- Prognatisme subnasal moderat.
- Ossos maxil·lars massius.
A la mandíbula:
- Mandíbules massives i robustes.
- Arc mandibular intern en “V” o “U” aguda.
- Arcada dental parabòlica sense diastema.
A les dents:
- Canines espatulades amb desgast perpendicular a la dent.
- Premolars i molars absolutament i relativament grans.
L’anatomia dels australopitecins mostra així mateix una sèrie d’adaptacions per a la bipedestació, tant en la part superior com en la inferior del cos. Entre les adaptacions de la part inferior s’inclouen les següents:
- L’ili (part de l’os coxal que forma la part anterior i lateral de la pelvis òssia) era molt més curt i ample que en els simis, el que permetia als músculs equilibrar el cos després de cada pas.
- La pelvis tenia forma còncava per allotjar els òrgans interns durant la postura dreta.
- La part alta dels membres inferiors formaven un angle cap a l’interior des de l’articulació del maluc, permetent així als genolls suportar millor el pes del cos en caminar dret. Al contrari, els membres inferiors dels simis estan col·locats gairebé en sentit vertical des del maluc, de manera que quan caminen drets el seu cos es balanceja cap als costats.
- Els dits dels peus eren més curts i menys flexibles que els dels simis, de manera que actuaven com a palanques per impulsar el cos a cada pas.
Per sobre de la pelvis també es van produir altres adaptacions. La columna australopitecina presentava una corba en «S» que disminuïa la longitud total del tors i li conferia rigidesa i equilibri quan es trobava dret; els simis, per contra, tenen una columna relativament recta. El crani australopitecí també presentava una adaptació important relacionada amb la bipedestació: el foramen magnum es trobava en una posició més avançada que en els simis, la qual cosa permetia al cap mantenir-se en equilibri sobre la columna dreta.
És clar que els australopitecins caminaven drets sobre el terra, però els paleoantropòlegs no tenen una opinió unànime sobre si passaven també una part important del seu temps en els arbres. Algunes característiques físiques com, per exemple, els dits corbs i llargs i els braços allargats, semblen confirmar que així era. Tanmateix, els dits, a diferència dels simis, potser no eren prou llargs com per permetre’ls balancejar-se de branca en branca.
La morfologia bàsica de l’australopithecus
Observem quines són les diferències anatòmiques més importants dels hominins respecte els pongins:
- El peu: s’ha especialitzat a estar dempeus, caminar i córrer. El dit gros ha perdut pràcticament l’oposició.
- Braços i mans: el braç és curt en relació amb les cames (els dos braços estirats = alçària total del cos). Els braços dels pongins toquen pràcticament a terra. Els hominins són els únics animals que poden recórrer còmodament distàncies llargues mentre porten objectes a les mans. La mà també és més curta que la dels pongins, i el polze, musculós i flexible, és totalment oposable als altres dits.
- Les cames dels hominins són les més llargues de tots els hominoides i el gran panxell n’és una característica distintiva. La musculatura glútia proporciona gran part de la força per a caminar costa amunt.
- La columna vertebral: té una peculiar corba a la regió lumbar, sense la qual el centre de gravetat del cos s’alteraria i tendiríem a caure cap enrere. És perpendicular al forat occipital; per això el cap es manté en equilibri dalt de la columna.
- El coll: és llarg i prim, ja que la postura vertical fa que el cap no necessiti poderosos músculs per a aguantar-se.
- La pelvis: té forma de conca i hi passa el centre de gravetat. Els ossos de la pelvis (sacre i ilíac) transmeten el pes del tronc a les cames. Proporciona acoblament als forts músculs que controlen les cames i incrementa la força de tota la musculatura que intervé en la posició erecta.
Aquestes diferències tenen un denominador comú: la postura vertical o dreta, és a dir, el bipedisme. Fa, doncs, uns 4 milions d’anys, pel cap baix, que els hominins són bípedes. Caminar sobre dues cames és una característica exclusiva dels homininis i és anterior al desenvolupament del cervell.
Ens cal preguntar ara: per a què els servia, a l’australopitec i als humans primitius, la postura erecta?
El bipedisme
Segons l’estat dels estudis actuals, en el període cronològic entre 7 i 5 milions d’anys, existeixen dubtes raonables per poder afirmar si havia aparegut la marxa erecta i si algun dels gèneres Sahelanthropus, Orrorin i Ardipithecus, o cap d’ells, era bípede. En canvi, del gènere Australopithecus es disposa de material fòssil suficient com per conèixer el seu tipus de locomoció. Més concretament, en els fòssils de l’espècie Australopithecus anamensis i, més encara, d’Australopithecus afarensis podem reconèixer les adaptacions a la locomoció bípeda, en fòssils que tenen fins a 4 milions d’anys d’antiguitat.
La consecució del bipedisme és un dels eixos centrals del camí cap a l’homo sapiens. Una concepció basada en la primacia del mascle té molt clara la funció i l’origen del bipedisme: som bípedes perquè som caçadors. La caça seria, doncs, el motor del desenvolupament humà; resumidament, aquest és l’argument:
La caça obliga a adoptar una postura erecta, tant per a veure les preses com per a fugir d’elles; i el caminar bípede deixa les mans lliures per a l’ús d’estris i la fabricació d’instruments, cosa que desenvolupa el cervell i redueix les canines, atès que no les hem de fer servir per atacar o defensar-nos (podem fer servir les mans amb pals) i, conseqüentment, la mandíbula també es redueix, possibilitant un augment de la grandària i la capacitat cerebrals.
Com podem criticar aquesta concepció androcèntrica?
Si el que ens va fer evolucionar hagués estat la caça d’animals grans, no s’entén la postura vertical del cos: tots els caçadors no humans del territori de l’Àfrica oriental on vivien els hominins eren quadrúpedes. El bipedisme no és pas una bona adaptació per a la caça: amb quatre potes es pot córrer molt més de pressa. Ni per la dentició (l’australopitec mai no hauria pogut matar a mossegades un animal gran), ni per les cames (en curses curtes no tenim res a fer amb la majoria d’animals), ni pels primitius còdols tallats, els hominins mai no han estat uns depredadors, i encara menys, mones assassines. En la transició d’animal a ésser humà, la caça no va tenir en absolut el paper que se li atribueix.
Si la caça no era, doncs, el seu sistema de subsistència bàsic, per a què li servien les eines que se li associen?
Quins avantatges tenia el bipedisme en un hàbitat no boscós? Des de Charles Darwin, s’han proposat nombroses i variades hipòtesis per explicar l’origen del bipedisme i totes elles intenten explicar per què durant el procés evolutiu es va seleccionar el bipedisme dels hominins. Algunes d’elles són:
- permet atènyer els fruits dels arbres baixos
- aixecar-se per damunt de les herbes de la sabana i tenir una millor visió que permetés localitzar les fonts d’aliment i alertar de possibles depredadors
- permet viatjar per la sabana i recórrer grans distàncies sense fer grans esforços;
- deixa les mans lliures per a agafar, llançar, aguantar i elaborar instruments lítics
- en termes del cost energètic, perquè el cos d’un bípede a determinades velocitats i distàncies és energèticament molt eficient
- dóna millors oportunitats d’apropar-se a un objectiu concret, la distància al qual es podia calcular amb precisió
- com adaptació tèrmica, perquè el cos d’un bípede exposa una menor superfície a la insolació i allunya el cap del terra
La hipòtesi de la termoregulació
Peter Wheeler[1] ha proposat una hipòtesi que relaciona la locomoció amb la vida en la sabana i els requisits de termoregulació que aquest hàbitat comporta. A les hores de màxima insolació, un bípede exposa una àrea molt més petita del seu cos a la radiació solar. En la part del cos més exposada, el cap, disposem de la protecció del cabell, mentre que la resta del cos hem desenvolupat la sudoració per evitar el sobreescalfament. A més, prop del terra l’aire està lleugerament més calent, mentre que a dos metres hi ha una més gran circulació d’aire. La simultània pèrdua de cabell en altres zones del cos va poder tenir com objectiu augmentar la sudoració per eliminar calor.
Segons Wheeler, el bipedisme reduiria la calor i el consum d’aigua en els hominins bípedes respecte als quadrúpedes. Aquesta adaptació a més permetria als hominins mantenir-se actius en les hores de màxima calor.
Un dels problemes sense resoldre d’aquesta proposta és la possibilitat que els primers hominins bípedes poguessin sorgir en ecosistemes més forestals, on la radiació solar no és determinant.
La hipòtesi de Lovejoy
Lovejoy[2] defensa que la locomoció bípeda d’Australopithecus afarensis va anar acompanyada per un conjunt d’adaptacions en el comportament. Segons aquest autor una de les innovacions evolutives fonamentals dels hominins fou l’aparició de la família nuclear: la monogàmia i el fet de tenir cura dels fills a càrrec dels progenitors. Segons aquesta hipòtesi, al mascle li correspondria proveir la família, així va aparèixer el bipedisme que permetria poder transportar aliments. Gràcies a aquest suplement energètic augmentava la capacitat d’alimentar a les cries i disminuïen els intervals entre naixements, i es podia criar a més d’un fill alhora. Però per tal que els mascles complissin aquest paper caldria l’existència de vincles més estables entre mascles i femelles per garantir que les cries són efectivament les que porten els seus gens. És a dir, Australopithecus afarensishauria de tenir un tipus d’organització familiar monògama.
Tanmateix, és difícil mantenir que Australopithecus afarensis era un hominí monògam atès que sembla que el dimorfisme sexual en aquesta espècie és molt marcat. El dimorfisme sexual, les diferències de mida entre mascles i femelles, és més gran en aquelles espècies en les quals els mascles han de competir entre ells per aparellar-se amb les femelles, en la majoria dels casos perquè els mascles intenten reproduir-se amb diverses femelles i impedeixen que altres mascles s’aparellin amb elles, assegurant d’aquesta forma la seva descendència. En aquestes espècies on els grups socials estan formats per un mascle i diverses femelles, la selecció natural afavoreix als mascles de mida superior. En els primats, les diferències en la dimensió corporal de mascles i femelles també venen acompanyades de grans diferències en la dimensió de les canines (les dels mascles són molt més gran que les de les femelles). Australopithecus afarensis presenta una combinació d’alt dimorfisme sexual corporal amb baix dimorfisme en la canina, i això fa difícil inferir el seu tipus d’organització familiar
Medi i estratègies de supervivència
L’australopitec és un hominí que viu en hàbitat obert (zones d’estepa o planes herbàcies), de postura erecta (no fa servir les mans per caminar), amb els peus adequats per a córrer i no per a grimpar, amb uns braços massa dèbils per a anar penjat per les branques i la columna vertebral més recta que la corbada dels quadrúpedes.
L’Africà fa 1,20 m, pesa uns 30 kg, té una capacitat cranial d’uns 450 cc i menja de tot (herbes, llangardaixos, ous, mamífers petits…). El robust fa 1,50, pesa uns 50 kg, la seva capacitat cranial supera els 500 cc i és bàsicament vegetarià.
«Els instruments no s’utilitzaren per caçar animals grans, ràpids i perillosos, sinó per a recol·lectar plantes, ous, mel, tèrmits, formigues i, probablement, petits animals excavadors. Els utensilis consistien en pals per a furgar o rebaixar la terra, roques per trencar núcules o altres fruits de closca dura, i diferents tipus de recipients. Les roques de tall fi s’utilitzaren potser també per tallar alguns dels aliments nous de la sabana, especialment rels i tubèrculs, durs i fibrosos, però prou voluminosos com per a fragmentar-los. Tot plegat és part d’un viratge general, en el que les mans i els estris substituïren les dents en algunes de les funcions.»
Nancy Tanner i Adrienne Zihlman, citat per R.E. Leakey: La formación de la humanidad. Serbal. Barcelona. 1981. pàg. 95
Si estava tan ben adaptat, per què va desaparèixer?
Australopithecus anamensis
Australopithecus anamensis és una espècie d’hominí de 4,2-3,9 milions d’anys d’antiguitat trobada a Kenya i descrita el 1995 per Meave Leakey. El nom d’aquesta espècie prové de la paraula tukana «anam» que significa llac i va ser elegida per la proximitat de Kanapoi al llac Turkana, i com a recordatori del gran llac que hi va haver en aquestes desertes zones en aquella època, el llac Lonyumun[3].
Els seus queixals tenien gruixuts esmalts, per la qual cosa es dedueix que no menjava només fulles i fruits, sinó aliments més durs. Va viure en un ambient forestal, però més obert que el dels seus presumptes avantpassats els Ardipithecus, i se sap que caminava dret. Presentaven un clar dimorfisme sexual en mida corporal. És possible que formessin comunitats de diversos mascles emparentats, en les quals cada un agruparia un petit harem de femelles.
L’equip de Tim White va trobar el desembre de 2005 restes d’aquesta espècie al lloc Asa Issie, al nord-est d’Etiòpia, a la vall mitjana del riu Awash. Datats amb entre 4,1 i 4,2 milions d’anys d’antiguitat, són un fèmur, fragments de maxil·lar i algunes dents, incloent l’ullal més gran trobat entre els hominins. Havien estat trobats a més uns altres 9 fòssils d’aquesta espècie, a Kanapoi, Kenya, la majoria el 1994; més uns altres 12 fòssils, principalment dents, trobades el 1988, a Allia Bay, Kenya i un fragment d’húmer que havia estat trobat el 1965 per Bryan Patterson i es va identificar posteriorment[4].
Encara que les dents i una mandíbula presenten encara similitud amb els simis, tenen, per primera vegada en l’evolució dels hominins, un esmalt gruixut (característica que comparteixen tots els Australopithecus, cosa que indica un canvi en el tipus d’alimentació i de nínxol ecològic, atès que han incorporat a la seva dieta aliments més durs[5]). El fragment d’húmer és semblant al dels humans i la part de tíbia trobada el 1994 per Kamoya Kimeu i el fèmur d’Asa Issie són la més antiga evidència del bipedisme erecte dels hominins. A partir de les característiques anatòmiques[6] dels ossos dels braços (llargs braços i les falanges corbades) i de les cames es dedueix que Australopithecusanamensis era, alhora, bípede i un poderós grimpador. El seu hàbitat, en comparació al dels homínids que el precedeixen, era un lloc més ideal per a l’evolució del bipedisme, amb regions menys poblades d’arbres i sabanes.
Segons alguns autors, Australopithecus anamensis seria l’avantpassat directe de l’Australopithecus afarensis (3,9 – 3 milions d’anys)[7]; seria un exemple d’anagènesi, és a dir, d’un canvi progressiu en lloc d’una bifurcació (cladogènesi). Segons Tim White l’Australopithecus anamensis mostra una clara descendencia del gènere Ardipithecus.[8]
Australopithecus afarensis: la Lucy
Australopithecus afarensis va ser un hominí que va viure entre els 3,9 i 3 milions d’anys abans del present. Era de contextura prima i gràcil, i es creu que va habitar només a Àfrica Oriental (Etiòpia, Tanzània i Kenya). És acceptat entre la majoria de la comunitat científica que pot ser un dels avantpassats del gènere Homo.
Va ser descobert el 24 de novembre de 1974 per Donald Johanson[9], Yves Coppens i Tim White a Hadar, Etiòpia. L’espècimen trobat en aquell moment va ser el mundialment conegut com «Lucy» (AL-288-1).
Aquest descobriment va destacar sobre els altres per moltes qualitats, especialment per ser l’Australopithecus més ben conservat a través de tants anys. Així va ser possible comprovar que la seva capacitat per caminar dret, com els humans actuals, va precedir el creixement del cervell. L’estudi de la seva dentadura va aclarir aspectes fonamentals sobre l’evolució dels hominins i va descobrir l’evolució simultània de gèneres, de manera que la línia Paranthropusse’n va apartar i en particular de la que va evolucionar cap a Homo.
Australopithecus afarensis és, com tot Australopithecus, un primat bípede dret, però diferent de nosaltres en diversos aspectes:
- capacitat cranial bastant menor: de 375 a 550 cm3. Això significa un cervell d’una mida propera a la tercera part de la humana actual mitjana.
- Australopithecus afarensis era un hominí amb freqüent activitat arbòria, ja sigui recollint fruits, dormint, jugant, etc.
- el seu pit no era en forma de barril, com el nostre, sinó que s’estrenyia agudament cap a dalt (forma de campana).
- el dimorfisme sexual era molt marcat, amb els mascles molt més corpulents que les femelles.
Pel que fa al seu sistema ossi és molt important recalcar que tenia el crani relativament petit, com els antropomorfs, encara que en comparació amb la mida del cos era relativament gran. A més, la seva cara era bastant gran i es projectava davant del crani (prognatisme), a causa de la mida de les dents.
Comparació de cranis d’Australopithecus afarensis i ximpanzé: a. Inclinació del front; b. Solc sobre el torus supraorbitari; c. Torus supraorbitari; d. Morfologia de la canina; e. Posició del foramen magnun
A diferència dels antropomorfs, els ullals d’Australopithecus afarensis són reduïts encara que es projecten lleugerament davant de la dent adjacent. Les incisives són grans (associades al règim frugívor). Els molars i premolars són de mida substancial, amb superfícies planes.
De la seva boca també es conclou que el paladar és molt similar al dels humans actuals perquè, encara que gran, forma una corba que no és parabòlica, ni de costats paral·lels, com en els grans simis.
La forma de la pelvis és important ja que se’n han realitzat estudis necessaris per determinar el caminar bípede dret dels Australopithecus; és de petita mida en comparació a la dels antropomorfs i presenta un canal de part, en la femella, més petit que l’actual i amb una notòria forma de ronyó. Les femelles de l’espècie no necessitaven tenir un gran canal de part, a causa de la petita mida del crani de les cries.
Segons Johanson i Tim White[10] l’origen dels humans primitius respondria a l’esquema següent: de l’australopithecus afarensis n’hauria sortit, per una banda l’australopithecus africanus i el robustus i, per l’altra, l’homo habilis. Segons altres autors, el crani més complert d’aquesta espècie, AL-444-2, trobat el 1992, presenta algunes característiques en el seu aparell masticatori i a la base del crani que el situarien en la línia dels Paranthropus, i qüestionen que aquesta espècie sigui l’avantpassada del gènere Homo.[11]
Però, fa aproximadament un milió d’anys, els australopitecs s’extingiren i només sobrevisqué una línia d’hominins: l’Homo habilis. El gènere Homo, doncs, degué començar la seva existència fa uns 4 milions d’anys, i durant 3 milions va coexistir amb l’australopitec. Ni l’Homo habilis ni l’australopitec es van moure d’Àfrica, i això ens planteja un seguit des qüestions:
- Per què va sobreviure i no s’extingí com l’australopitec?
- Per què el cervell de l’Homo habilis va multiplicar per dos la capacitat de la de l’australopitec?
- Què té, què fa (o com ho fa) el gènere Homo que no tingués o fes el gènere australopitec?
- Què tenia l’Àfrica oriental que va ser també el bressol de l’Homo habilis?
Museo Nacional de Antropología de México Lucy Cleveland Natural History Museum
Reconstrucció de la cara de la nena Selam[12], 100 000 anys més antiga que la Lucy. Fou descoberta per l’antropòleg etíop Zeresenay Alemseged l’any 2000 a deu quilòmetres de distancia de la Lucy.
Reproducció d’A.afarensis a Cosmocaixa, Barcelona
El 24 de novembre de 1974, Donald Johanson i el seu equip van trobar en el riu Awash, a la regió d’Afar (a Etiòpia), un esquelet d’homínid. Es va poder recuperar el 40 % dels seus ossos, que tenien una antiguitat d’uns 3 milions d’anys. A aquest esquelet, que correspon a un australopitec femella, li van posar el nom de Lucy per la cançó dels Beatles Lucy in the sky with Diamonds, que sonava a la ràdio del campament. Les restes de Lucy van ser trobades al lloc on habitava la tribu Afar, per això el nom afarensis.
Es tracta d’una femella adulta, d’uns 20 anys, que caminava bípeda i feia 1,20 d’alçada. Els seus braços, en relació amb les seves cames, són una mica més llargs que els dels humans actuals, però no tant com els dels pongins. La mà ja té un polze que es pot oposar a l’índex, fet que li permet la «prensió de precisió» (pot enfilar una agulla, cosa que no pot fer un ximpanzé).
La Lucy no és un ésser humà, però tampoc no és ni un mico, ni un ximpanzé…. És un hominini del gènere australopitec: pertany a l’espècie Australopithecus afarensis.
De fet, el terme australopitec, que vol dir «mona del sud», no és correcte, atès que els australopitecs no són mones. Però és que les primeres restes d’un ésser similar a la Lucy foren trobades el 1924 pel sud-Africà Raymond Dart a Taung, en una cova de l’actual Botswana, i li va donar el nom d’australopitec, pensant que es tractava d’un simi evolucionat; i el primer nom que es dóna a una espècie acabada de descobrir és d’obligatòria imposició per a tots els altres membres d’aquesta espècie que es trobin més tard.
Les petjades de Laetoli
Les petjades de Laetoli es van formar sobre cendres molt fines expulsades pel volcà Sadimán fa 3,6 milions d’anys. Després de l’erupció va ploure (hi ha marques de gotes de pluja) i el terra es va humitejar. Els hominins i altres animals van passar i van deixar el seu rastre. El sol va sortir i va assecar ràpidament els rastres i, donat l’alt contingut en carbonats, es va endurir com el ciment. Van caure altres capes de cendres fins a cobrir uns 16-20 centímetres i les empremtes van quedar enterrades i conservades.
Presenten un patró morfològic totalment humà. El cop de taló està molt marcat, el dit gros està completament alineat amb la resta dels dits i l’arc plantar medial està ben desenvolupat. Les relacions espacials entre les empremtes presenten un patró sorprenentment humà (distància entre les trepitjades, orientació de les mateixes, etc.).
Es distingeixen tres rastres de trepitjades. En els anys setanta del segle XX es van descobrir rastres de dos individus desplaçant-se cap al nord. Són dos rastres paral·lels, separats entre si 25 centímetres, massa pròxims per a dues persones caminant una al costat de l’altra. És possible que seguissin el mateix camí però que no es produïssin de forma simultània, ja que les condicions d’ambdós rastres són una mica diferents. El primer dels rastres (conegut com a rastre G-1), el de peus més petits, presenta els contorns nítids, com si s’haguessin format sobre una superfície ferma i compacta (humida), mentre el rastre del peu major (rastre G-2) té els contorns més difusos i engrandits, com si s’haguessin produït més aviat sobre una superfície seca i polsosa. El rastre G-1 sembla detenir-se a meitat de camí, donar mitja volta cap a l’esquerra, i tornar a reprendre el camí inicial.
No va ser fins després quan els investigadors es van adonar que el rastre situat més a l’est (rastre 2) representava empremtes superposades de dos individus: G-2 la més profunda i G-3 la més superficial.
A partir de les empremtes s’han fet estimacions de l’estatura, però amb diversos pressupòsits de partida:
- els hominins de Laetoli tenen les mateixes proporcions del peu que els humans moderns.
- els individus representats són adults.
- les trepitjades són bons indicadors de la mida del peu.
Usant diverses poblacions actuals, el rang obtingut per a la major de les petjades és d’1,34 a 1,56 metres i per a la de menor mida d’1,15 a 1,34 metres. Aquestes estimacions són consistents amb les obtingudes a partir de restes de l’esquelet postcranial d’Australopithecus afarensis, el candidat més adequat com a responsable d’aquestes petjades.
Aquest rastre permet estudiar el tipus de locomoció dels hominins que el van produir. A les empremtes de Laetoli, com en les d’un humà actual, la transmissió del pes comença al taló, segueix al llarg de la vora exterior i acaba al dit gros, com es podria esperar en un bípede.
En aquest jaciment (Laetoli) s’han trobat restes fòssils d’Australopithecus afarensis, la qual cosa permet especular amb certa seguretat que individus d’aquesta espècie són els autors de les empremtes.
Australopithecus africanus
Australopithecus africanus és una espècie d’hominí fòssil de Sud-àfrica, datat en uns 5 milions d’anys. La història d’Australopithecus africanus com a espècie reconeguda data de 1924, quan va ser descobert per Raymond Dart a Taung, Sud-àfrica, el crani d’un nen conegut com el Nen de Taung. Però no va ser tingut en compte fins 30 anys després.
El crani més complet d’Australopithecus africanus conegut com Mrs. Ples, de 2,1 milions d’anys d’antiguitat i una capacitat cranial d’uns 485 cm3. Transvaal Museum. Fou descobert el 18 d’abril del 1947 per Robert Broom i John T. Robinson a la regió de Sterkfontein, a uns 70 k,m al sud-oest de Pretòria.
D’acord amb els estudis dentals realitzats, Australopithecus africanus procedeix d’Australopithecus afarensis i fins no fa molt es creia que obria una línia evolutiva cap al gènere Paranthropus, i una altra línia, que també procedeix d’Australopithecus afarensis, va estar representada per Homo habilis i va culminar en l’Homo sapiens.
Entre les característiques es remarca la forma de caminar bípeda, comuna a tots els hominins. Tenien activitat arbòria en forma activa. El seu pes mitjà oscil·lava entre els 33 i 67 quilograms, amb una estatura d’1,50 metres d’alt.
Les seves característiques òssies són de gran importància per diferenciar-se de la resta d’Australopithecus: la seva capacitat craniana és de 480 a 520 cm3. La seva caixa cranial és més alta i arrodonida que la dels antropomorfs, i el seu esquelet postcranial similar al de l’Australopithecus afarensis.
La seva cara, des d’un punt de vista general, és més curta. Presenta menor prognatisme (la mandíbula està avançada respecte el maxil·lar), unit a una menor mida de peces dentals. Aquestes no es diferencien gaire de les de l’humà. Les canines eren petites, sense sobresortir com en els antropomorfs, i les incisives eren tan petites com les nostres. També és important esmentar que desapareix la diastema (separació entre incisives), o és molt rara la seva existència. Per tant, notem una reducció de canines i incisives i hi ha un major èmfasi de la masticació en la resta de la dentadura.
Australopithecus bahrelghazali
Australopithecus bahrelghazali és una espècie fòssil d’hominí trobada el 1995 per Michel Brunet a Bahr el-Ghazal, Txad, representat per una mandíbula amb set dents de 3,5 a 3 milions d’anys d’antiguitat. Va ser sobrenomenat Abel. És la primera evidència de fòssils d’hominins a l’oest de la vall del Rift. Es considera que representa una línia d’Australopithecus diferent de la que va evolucionar cap a Homo.
KT12/H1. Abel. Australopithecus baherghazali
Dentició:
- Esmalt més gruixut que Ramidus.
- Ús intens de les dents anteriors.
- Incisives i canines amb corones altes i llargues arrels.
- Premolars amb tres arrels. El tercer premolar té dues cúspides i tres arrels, mentre que el quart premolar està molaritzat. Els tercers premolars superiors presenten corona asimètrica i tres arrels. Precisament el fet de tenir premolars amb tres arrels el diferencien clarament d’Afarensis, el qual presenta una o dues arrels a P3 i tres a P4. A més la forma de la mandíbula és diferent en ambdues espècies. P3 és més asimètric que oval, com en els Paranthropus.
Pel que fa al seu mode de vida, el seu hàbitat era bastant obert dominat per pastures tropicals. L’anàlisi d’isòtops de carboni presents en els ossos revela una dieta basada en pastures[13] (Julia Llegeix-Thorp et al, 2012).
Amb tan pocs fòssils, és impossible determinar la posició filogenètica de l’espècie.
Spoor[14] suggereix una relació amb Kenyanthropus, a causa de la similitud del seu ortognatisme, però Josephine C.A. Joordens pensa que Australopithecus afarensis va poder arribar a la conca del Txad aprofitant un episodi humit.
East Side Story fou una hipòtesi proposada per Yves Coppens[15] el 1994 que intenta explicar l’aparició dels homínids bípedes. Aquesta hipòtesi es basa en els següents fets:
- la zona oest de l’Àfrica compresa entre l’oceà Atlàntic i la vall del Rift està poblada per ecosistemes selvàtics, mentre que a la zona a l’est de la vall del Rift fins a l’oceà Índic predominen els ecosistemes de sabana.
- els homínids bípedes més antics s’han trobat a l’est de la vall del Rift.
Segons l’esmentat autor, durant el Miocè, un gran cinturó de selva s’estenia des de l’Atlàntic fins a l’Índic. La formació de la vall del Rift[16] (finals del Miocè) a causa de processos tectònics, amb els consegüents canvis en el relleu, va aixecar grans barreres muntanyoses que han actuat des de llavors com a pantalla orogràfica; així els vents carregats d’humitat provinents de l’Atlàntic no poden superar aquesta pantalla i a l’est de la mateixa es redueix la precipitació. Els grans simis van quedar aïllats en poblacions occidentals (avantpassats de goril·les i ximpanzés) i poblacions orientals. Aquestes es van haver d’adaptar al nou ecosistema més sec (sabana, bosc obert) el que les va empènyer a abandonar la vida arborícola per esdevenir habitants del terra. La millor solució per a un homínid que es veu obligat a desplaçar-se pel terra per buscar aliment i refugi en arbres veïns seria el bipedisme.
La hipòtesi ha estat criticada després de la troballa dels fòssils d’Australopithecusbahrelghazali al Txad, a l’oest de la vall del Rift, així com de restes d’homínids suposadament bípedes no associats a ecosistemes oberts (Orrorin tugenensis, Sahelanthropus tchadensis, Ardipithecus)[17].
Australopithecus garhi
Australopithecus garhi és una espècie d’hominí els fòssils del qual es van descobrir el 1996 per un grup d’investigadors liderat pel paleontòleg etíop Berhane Asfaw i l’antropòleg nord-americà Tim White[18].
Inicialment es va creure que era la baula perduda entre els gèneres Australopithecus i Homo, i per tant un avantpassat de la nostra espècie. Tanmateix Australopithecus garhi és més avançat que qualsevol altre australopitec, i una espècie contemporània (o gairebé) de les espècies ancestrals d’Homo, per tant no és probable que sigui un avantpassat humà. Les restes trobades procedeixen d’un lapse temporal amb un escàs registre fòssil, entre 2 i 3 milions d’anys. Tim White va ser el científic que va trobar el primer dels fòssils d’Australopithecus garhi el 1996 a prop de la ciutat de Bouri, al riu Awash mitjà de la Depressió d’Afar, a Etiòpia. L’espècie va ser confirmada i va ser establerta com Australopithecus garhi el 20 de novembre de 1997 pel paleoantropòleg etíop Yohannes Haile-Selassie. El nom de l’espècie “garhi” significa “sorpresa” en la llengua local Afar.
Els trets d’Australopithecus garhi són una mica diferents dels típicament observats en Australopithecus afarensis i Australopithecus africanus. Un exemple de les diferències pot veure’s en comparar el maxil·lar trobat a Hadar, Etiòpia (Australopithecus afarensis) amb l’espècimen de Bouri d’Australopithecus garhi. La capacitat cranial d’Australopithecusgarhi és de 450 cm3, semblant a la d’altres australopitecs. La mandíbula trobada per Asfaw té una morfologia general compatible amb la mateixa espècie, encara que és possible que pertanyi a altres espècies d’hominins trobades en els mateixos dipòsits. Els estudis fets als premolars i molars mostren certa similitud amb els de Paranthropus boisei, que és més gran que en altres formes gràcils d’australopitecs. S’ha suggerit que si Australopithecus garhi és un avantpassat d’Homo la morfologia maxil·lar hauria d’haver hagut d’evolucionar ràpidament, en amb prou feines 200.000 o 300.000 anys.
Els pocs artefactes primitius de pedra descoberts amb els fòssils d’Australopithecus garhi, datats en 2,5-2,6 milions d’anys, recorden remotament la tecnologia Olduvaiana. Aquestes eines són més antigues que les d’Homo habilis, que és considerat un possible ascendent directe d’hominins més moderns. Així mateix, a Bouri es van trobar uns 3.000 artefactes toscos de pedra d’una edat estimada en 2,5 milions d’anys. Aquestes antigues eines (còdols, palets i ascles) mostren l’ús de diverses tècniques que es poden observar en les indústries més tardanes Olduvaiana i Acheuliana.
Junt amb els fòssils hominins s’han trobat també ossos d’antílops que mostren marques de tall curvilínies realitzades amb instruments de pedra. Alguns dels ossos d’antílop mostren evidències d’haver estat oberts a cops amb martells de pedra. Això indicaria que aquests hominins eren capaços d’adquirir grans quantitats de recursos alimentaris d’alta qualitat (carn i greix de la medul·la òssia).
Des del punt de vista de la tecnologia i de la dieta, Australopithecus garhi representa una primera etapa cap a l’aparició del gènere Homo. Abans del descobriment d’Australopithecus garhi, l’antecedent més antic d’un hominí capaç d’utilitzar un instrument de pedra estava representat pel gènere Homo, amb una antiguitat de 2,3 milions d’anys i trobat també a la mateixa regió. La relació entre Australopithecusafricanus i el gènere Homo era molt difícil de desxifrar a causa del buit existent en el registre fòssil de l’Àfrica de l’est entre els 2 i 3 milions d’anys. Australopithecus garhi ve a omplir aquest buit i a demostrar que probablement l’ús d’instruments i el canvi de dieta es va produir abans de l’augment de la capacitat cranial.
Australopithecus sediba
El lloc de Malapa, a Sud-àfrica, ha donat el conjunt més ric de fòssils d’homínids primerencs al continent d’Àfrica. Les restes fòssils d’Australopithecus sediba van ser descobertes a l’agost del 2008[19], i l’espècie va ser nomenada el 2010 i s’ha datat amb una antiguitat de 1,9 a 1,98 milions d’anys. Seria, doncs, l’australopitecí més modern.
Al juliol del 2012, científics de l’Institut Wits sobre l’Evolució Humana, de la Universitat de Witwatersrand, van anunciar el descobriment d’un esquelet d’Australopithecus sediba en una roca d’un metre de diàmetre que havia estat ignorada en un laboratori, fins que un tècnic va percebre una dent a la superfície. La peça va ser sotmesa a un escàner que va revelar el que sembla ser un fèmur complet, costelles, vèrtebres i altres parts importants de les extremitats[20].
Dent d’un homínid incrustat en una roca que conté parts significatives d’un esquelet d’un avantpassat humà primerenc. Podria formar part de “Karabo”, l’esquelet d’Australopithecus sediba descobert a Malapa.
Comparteix amb Australopithecus africanus nombroses característiques a la volta del crani, l’esquelet facial, la mandíbula i les dents. Però altres característiques (dents més petites, pòmuls menys pronunciats, nas prominent, cames més llargues, trets de la pelvis) l’apropen a Homo. No obstant això, basant-se en alguns trets entre els quals destaca la petita grandària del cervell, s’ha inclòs dins del gènere Australopithecus.
L’esquelet postcranial, és similar al d’altres australopitecins: el seu cos era petit, els seus membres superiors relativament llargs, i retenia característiques primitives en els membres superiors i inferiors. Difereix d’altres australopitecins i s’apropa a Homoen algunes formes que anticipen la reorganització de la pelvis i membres inferiors en l’Homo erectus. Cal destacar que aquesta reorganització de la pelvis es dóna abans que es produeixi l’augment de mida del cervell del fetus, que es produirà posteriorment en la filogènia humana.
Comparació dels còccix d’Australopithecus afarensis (AL 288-1), Australopithecus africanus (Sts 14), Australopithecus sediba (MH1), y Homo erectus (KNM-WT 15000).
Un dels esquelets, un jove d’uns 12-13 anys, està representat per un crani parcial, un fragment de mandíbula i part de l’esquelet postcranial. La seva grandària cerebral (420 cc) es considera que arribava al 95% de la mida adult.
Una altre esquelet, el d’una femella adulta, està representat per una dent maxil·lar, una mandíbula parcial, i part de l’esquelet postcranial. De l’anàlisi de les mans, amb dits curts i polze relativament llarg, capaços de realitzar la pinça de precisió, es pot inferir la capacitat per produir eines lítiques.
Els llargs braços i l’estructura dels peus suggereixen capacitat de grimpar. Es dedueix que aquesta espècie gaudia d’una àmplia varietat de comportaments i podia explotar diversos entorns.
S’estima que l’alçada màxima de l’espècie era 1,30 m.
De l’estudi d’aquestes restes se’n poden descriure les següents característiques:
Trets primitius: Tòrax superior estret i cònic. Aquesta morfologia limita el balanceig dels braços en caminar; braços llargs i simiescos i escàpula adaptats a la trepa i fins i tot a la suspensió; peu similar al del ximpanzé, encara que amb un turmell com el d’Homo, i el taló encara més primitiu que el d’Australopithecus afarensis.
Trets derivats en la línia de sapiens: curvatura de l’esquena baixa; columna flexible; mateix nombre de vèrtebres lumbars que Homo; canell, mà i dits adaptats a la prensió de precisió. Van poder fabricar eines sense perdre la capacitat d’escalada.
Dents: boca semblant a la humana; canines petites. Tot i que hi tenen una relació, són diferents als d’Australopithecus africanus en grandària, forma i trajectòria ontogenètica i això garanteix la separació filogenètica amb aquesta espècie. Les dents d’Australopithecus sediba (i també els d’Australopithecu africanus) estan més relacionats amb els d’Homo que les dents dels austalopitecins de l’est d’Àfrica. Suggereixen que Australopithecus sediba no és un descendent d’Australopithecus afarensis.
Australopithecus sediba. Restes corresponents a las extremitats superiors
Semblen possibles les següents interpretacions:
- Australopithecus sediba probablement és descendent d’ Australopithecus africanus, i sembla l’australopitec més proper a Homo.
- Té trets craneodentals i patrons de locomoció més avançats que Sterkfontein STW 53, Olduvai OH 62 i Koobi-Fora KNM-ER 3735, més o menys contemporanis en el temps i assignats a Homo habilis, amb ell que aquesta assignació esdevé incerta.
- El seu taló, més primitiu que el d’Australopithecus afarensis, fa pensar que forma part d’un llinatge diferent al representat per Australopithecus afarensis i Australopithecus africanus.
- Australopithecus sediba podria estar en el camí evolutiu cap a Homo ergaster. Aquesta alternativa és defensada, entre d’altres, per Chris Stringer[21].
- Homo habilis constituiria una branca aïllada i sense descendència de l’arbre d’Homo.
- Podria representar un grup paral·lel que va persistir durant algun temps després de la primera aparició d’Homo. Aquesta sembla la hipòtesi més probable, defensada entre d’altres per Donald Johanson.
En tot cas, sediba és una prova més que la confusa transició entre Australopithecus i Homo es va produir en forma de mosaic, amb canvis en la pelvis, augment de la distància de les línies temporals i reducció de la constricció postorbital abans que augmentés l’encefalització.
El dimorfisme sexual en sediba sembla similar al dels humans moderns.
La seva alimentació incloïa escorces toves, fulles, fruits i altres productes vegetals dels ambients boscosos i tancats, fins i tot durs, una alimentació generalista i diversa molt diferent a la de la resta dels australopitecins i primers Homo.
The Discovery of Australopithecus sediba https://www.youtube.com/watch?v=5YEiJVQdI-Q
https://www.youtube.com/watch?v=FwpQCjeIcy4
Kenyanthropus platyops
Rèplica del crani KNM-WT-4000, Museu en Brno
Kenyanthropus platyops (platyops = cara plana) és el nom proposat per a una espècie d’hominí de la qual se’n va trobar un crani fòssil el 1999. La troballa va ser feta per un equip dirigit per Meave Leakey i la seva filla Louise, a Kenya, en una regió semidesèrtica propera al llac Turkana[22]. El fòssil data d’uns 3,5 milions d’anys. Té característiques molt interessants, ja que té trets facials plans i delicats, i dents relativament petites. Però no mostra les adaptacions masticatòries dels Paranthropus, ja que no té cresta sagital i les dents són petites. Tampoc té la morfologia facial dels australopitecins, i no pot ser inclòs en el gènere Homo perquè té una capacitat encefàlica de només 400-500 cc.
Els científics han proposat definir el gènere Kenyanthropus, doncs el crani trobat mostra una mescla equilibrada de trets d’Australopithecus i Homo, s’assembla a «KNM-ER 1470», un altre fòssil trobat fa uns anys en aquesta regió, classificat com Homorudolfensis, i amb una datació molt menor, d’uns 1,8 milions d’anys. No obstant això, ja que l’espècimen tipus (KNM-WT 4000) està terriblement distorsionat, hi ha autors que no ho consideren ni un gènere ni una espècie vàlida i que podria tractar-se simplement d’un espècimen d’Australopithecus afarensis o fins i tot d’alguna espècie d’Homo, tal com Homo rudolfensis.
[1] Peter Wheeler (1993). «Human ancestors walked tall, stayed cool». Natural History, 102: 65-67
[2] Lovejoy, C.O. (1981). «The origin of man». Science 211: 341-350
[3] Leakey, Meave G., Feibel, C.S., McDougall, I., Walker, A. (1995): «New four-million-year-old hominid species from Kanapoi and allia Bay, Kenya.», Nature 376: 565-571
[4] Fredrick K. Manthi, J. Michael Plavcan i Carol V. Ward: « New hominin fossils from Kanapoi, Kenya, and the mosaic evolution of canine teeth in early hominins». S Afr J Sci. 2012;108(3/4), Art. #724, 9 pàgs. En línia a http://www.sajs.co.za/sites/default/files/publications/pdf/724-7328-7-PB.pdf
[5] Reconstruint la dieta de l’homínid africà ‘Australopithecus anamensis’. Notícies Universitat de Barcelona. Recerca. 9 juliol del 2012. En línia a http://www.ub.edu/web/ub/ca/menu_eines/noticies/2012/07/024.html. Ferran Estebaranz, Jordi Galbany, Laura M Martínez, Daniel Turbón & Alejandro Pérez-Pérez.«Buccal dental microwear analyses support greater specialization in consumption of hard foodstuffs for Australopithecus anamensis». Journal of Anthropological Sciences Vol. 90 (2012), pp. 1-24.
[6]C.V. Ward, M.G. Leakey, A. Walker: « Morphology of Australopithecus anamensis from Kanapoi and Allia Bay, Kenya». Journal of Human Evolution. Volum 41, nº 4. Octubre 2001. Pages 255–368.
[7] William H. Kimbel,Charles A. Lockwood,Carol V. Ward,Meave G. Leakey,Yoel Rak,Donald C. Johanson:« Was Australopithecus anamensis ancestral to A. afarensis? A case of anagenesis in the hominin fossil record». Journal of Human Evolution, Volum 51, nº 2, agost del 2006, Pàgs. 134–152.
[8] White, T.D, WoldeGabriel, G., Asfaw,B., Ambrose, S., Beyene, Y., Bernor, R.L., Boisserie, J.-R., Currie, B., Gilbert, H., Haile-Selassie, Y., Hart, W.K., Hlusko, L.J., Howell, F.C., Kono, R.T., Lehmann, T., Louchart, A., Lovejoy, C.O., Renne, P.R., Saegusa, H., Vrba, E.S., Wesselman, H., Suwa, G.: «Asa Issie, Aramis and the origin of Australopithecus.»Nature 440, 13 abril del 2006, pàgs.883-889. En línia a http://www.nature.com/nature/journal/v440/n7086/abs/nature04629.html
[9] Johanson, D.C. i Taieb, M.: «Plio-Pleistocene hominid discoveries in Hadar, Ethiopia». Nature 260, 25 de març del 1976, : 293-297. En línia a http://www.readcube.com/articles/10.1038/260293a0
[10] Johanson, D.C., White, T.D. «A systematic assessment of early African homninids». Science 203, nº 4378, 26 de gener de 1979, pàgs. 321-330.
[11] Arsuaga, Juan Luis i Martínez, Ignacio (2009). La especie elegida : la larga marcha de la evolución humana. Barcelona. Círculo de Lectores. 2ª impr. 360 pàgs.
[12] «Aquí es descriu un esquelet infantil ben conservat de 3,3 milions d’anys d’un Australopithecus afarensis descobert a l’àrea de recerca de Dikika, a Etiòpia. El crani d’una presumpta femella de tres anys mostra que la majoria de característiques de diagnòstic de les espècies són evidents fins i tot en aquesta fase inicial de desenvolupament. La troballa inclou molts elements d’un esquelet prèviament desconegudes en un registre d’homínids del Pliocè, incloent-hi un os hioide que té la morfologia típica dels simis africans. El peu i altres evidències de l’extremitat inferior proporcionen proves clares d’una locomoció bípeda, però l’escàpula típica de goril·la i les falanges llargues i corbes, plantegen noves preguntes sobre la importància del comportament arborícola en el repertori locomotor de l’Australopithecus afarensis».Alemseged, Zeresenay. «A juvenile early hominin skeleton from Dikika, Ethiopia». Nature, vol. 443, 7109, 21 de setembre del 2006, pàgs. 296–301. En línia a http://www.nature.com/nature/journal/v443/n7109/full/nature05047.html
[13] Matt Sponheimer, Julia A. Lee-Thorp, Kaye E. Redd (2013). Early Hominin Paleoecology. Boulder. University Press of Colorado. 368 pàgs.
[14] Meave G. Leakey, Fred Spoor, Frank H. Brown, Patrick N. Gathogo, Christopher Kiarie, Louise N. Leakey i Ian McDougall: «New hominin genus from eastern Africa shows diverse middle Pliocene lineages». Nature, 410, 22 de març del 2001, pàgs.433–40
[15] Coppens, Yves: «East Side Story: the origin of Humankind». Scientific American 270, nº 5, maig 1994, pàgs. 88-95
[16] Veure Gran Vall del Rift a http://ca.wikipedia.org/wiki/Gran_Vall_del_Rift
[17] El 1995 Michel Brunet va publicar la troballa d’un fragment anterior de mandíbula amb set dents a la regió Koro Toro del Txad, a 2.500 km de la vall del Rift, d’una antiguitat de 3,5 a 3 milions d’anys, i el va assignar a l’espècie Australopithecus bahrelgazhali. Brunet, M., Beauvilain, A., Coppens, Y., Heintz, E., Moutaye, A.H.E. i Pilbeam, D.: « Australopithecus bahrelgazhali, une nouvelle espèce d’hominidé ancien de la région de Koro Toro (Tchad). Comptes rendus de l’Académie des Sciences de Paris, série IIA, Earth and Planetary Science 322: 907-913
[18] Asfaw, Berhane; White, Tim; Lovejoy, Owen; Latimer, Bruce; Simpson, Scott y Suwa, Gen (1999). «Australopithecus garhi: a new species of early hominid from Ethiopia». Science Vol. 284, nº 5414, 23 d’abril del 1999, pàgs. 629-635. En línea a http://www.indiana.edu/~origins/teach/P314/Bouri.pdf
[19] L. R. Berger: «The Mosaic Nature of Australopithecus sediba». Science, 12 Abril 2013, Vol. 340 no. 6129 pp. 163-165. Veure també els textos reunits a http://www.sciencemag.org/site/extra/sediba/index.xhtml
[20] University of the Witwatersrand. «Early human ancestor, Australopithecus sediba, fossils discovered in rock.» ScienceDaily., 12 July 2012. En línia a http://www.sciencedaily.com/releases/2012/07/120712162744.htm
[21] Chris Stringer (2011). The Origin of Our Species. Londres: Allen Lane. 352 pàgs.
[22] Leakey, M.G., Spoor, F., Brown, F.H., Gathogo, P.N., Kiarie, C., Leakey, L.N., i McDougall, I.: «New hominin genus from eastern Africa shows diverse middle Pliocene lineages». Nature 410, 22 març del 2001, pàgs. 433-440.
Joan Campàs Aura digital
Curs: Orígens de l’Art i evolució humana: l’homo significans
Estudis d’Arts i Humanitats de la UOC
Materials del curs en format .pdf
1. Una visió de conjunt de l’art paleolític
2. El paleolític: cronologia i referències etnoarqueològiques
3. Evolució tecnològica en el Paleolític. Referències etnoarqueològiques de l’hàbitat
4. La construcció del coneixement de l’art del Paleolític
5. El paradigma d’Henri Breuil i la seva crisi
6. L’estructuralisme de Leroi-Gourhan i Annette Laming-Emperaire
7. Del xamanisme a l’art com expressió de l’organització social
8. El procés evolutiu: Darwin i la selecció natural
9. Darwin i la selecció sexual. El cicle menstrual
10. La selecció sexual: la competència espermàtica
11. La selecció sexual: l’orgasme femení
12. Sobre els orígens de l’art: les coalicions de dones pintades
13. Una perspectiva ecològica de l’evolució humana
14. Els primers hominoides
15. Els homínids: els preaustralopitecins
16. Els hominins: els australopitecins
17. Paranthropus
18. Els primers Homo: Homo habilis
19. El poblament d’Euràsia: l’homo ergaster/erectus
20. Homo antecessor, rhodesiensis, heidelbergensis i floresiensis
21. L’Homo neanderthalensis
22. L’Homo sapiens
23. Sobre el concepte d’art
24. Les primeres expressions artístiques
25. Sobre els orígens de l’art del Plistocè
26. L’art rupestre del Plistocè
27. Temes i distribució de l’art parietal
28. L’art moble del Plistocè
29. Temes de l’art mobiliari
30. Art mobiliari del Paleolític Superior. Interpretacions
31. Sobre els orígens del llenguatge