18. Els primers Homo: Homo habilis
1 desembre, 2014No se sap amb certesa quina espècie va originar els primers membres del gènere Homo; s’han proposat Australopithecus africanus, Australopithecus afarensis i Australopithecusgarhi, però no hi ha un acord general. També s’ha suggerit que Kenyanthropus platyopsva poder ser l’avantpassat dels primers Homo.
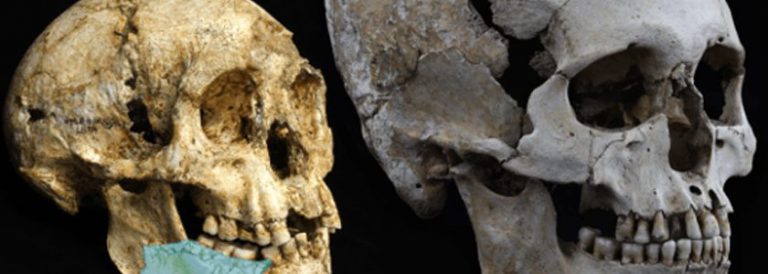
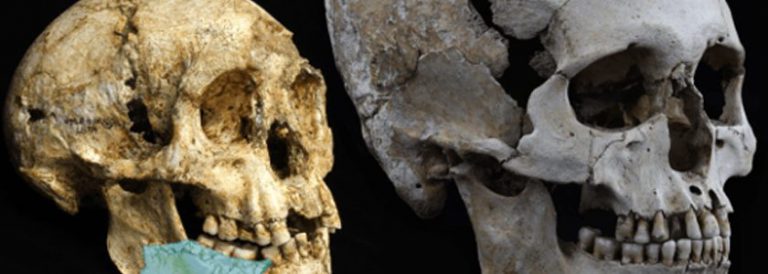
El millor candidat a ser l’avantpassat del gènere Homo és l’Australopithecus africanus, però aquest presenta una lleugera megadòncia que el diferencia d’Homo, el qual té les dents, especialment,les molars, més petites. Sí que sembla que es pot afirmar que els humans sorgeixen a partir d’un australopitec gràcil, similar a l’africanus.
Clàssicament es consideren com a pertanyent al gènere Homo els hominins capaços d’elaborar eines de pedra. No obstant això, aquesta visió ha estat posada en dubte en els últims anys; per exemple, s’ha suggerit que Australopithecus ghari, fa 2,5 milions d’anys, va ser capaç de fabricar eines. Les primeres eines eren molt simples i s’enquadren en la indústria lítica coneguda com Olduvaiana o Mode Tècnic 1. Les més antigues procedeixen de la regió d’Afar (Etiòpia) i la seva antiguitat s’estima en uns 2,6 milions d’anys, però no existeixen fòssils d’hominins associats a ells.
Els fòssils més antics que amb tota seguretat es poden atribuir al nostre gènere provenen d’Hadar[1] (Etiòpia). L’equip de Wiliam Kimbel i Donald Johanson[2] van trobar un maxil·lar molt complert (AL-666-1) amb una antiguitat d’uns 2,4 milions d’anys, associat a una vintena d’eines lítiques[3].
D’aquesta fase s’han descrit dues espècies, Homo habilis i Homo rudolfensis, que van habitar Àfrica Oriental entre 2,5 i 1,8 milions d’anys. El volum cranial d’aquestes espècies oscil·la entre 650 i 800 cm3.
L’Homo habilis
Cap el 1950 es deia que l’australopitec africà era l’avantpassat de l’homo sapiens. Però Louis Leakey s’hi va oposar: no hi havia prou temps entre l’australopitec i l’homo erectusper tal que el primer fos l’avantpassat del segon.
El 1964 va descobrir a Olduvai (nord de Tanzània) fragments cranians que segons ell pertanyien a un homínid més avançat que l’australopitec, i li va donar el nom d’Homo habilis (nom incorrecte perquè l’australopitec i, fins i tot, els ximpanzés, utilitzen eines); tenia una antiguitat d’uns 1,75 milions d’anys, una capacitat craniana de 680 cc, una mandíbula amb una filera de dents amb la típica forma d’U humana, i restes d’una mà i d’un peu, que segons John Napier mostraven que aquest humà era bípede i tenia capacitat per a fabricar estris (pedres treballades i pedres en cercle per recolzar-hi una cabana de branques). Associaven, doncs, dues característiques pròpies dels humans: desenvolupament encefàlic i elaboració de tecnologia lítica[4].
Posteriorment, a Olduvai s’han trobat més fòssils com els cranis OH 24 i OH 13, que abasten un període de 1,8 a 1,6 milions d’anys. S’han trobat fòssils d’Homo habilis a la vora oriental del llac Turkana (Kènia), entre els que sobresurt el crani KNM-ER 1813, així com en el jaciment sudafricà de Sterkfontenin amb una antiguitat de 1,8 milions d’anys, el crani STW 53.
KNM-ER 1813. http://humanorigins.si.edu/evidence/human-fossils/fossils/knm-er-1813
STW 53. http://www.scielo.org.za/scielo.php?pid=S0038-23532008000600015&script=sci_arttext
Crani i dentició d’Australopitec i d’Homo
Crani d’Homo habilis, OH 24 o “Twiggy”. Rèplica Original. Gorja d’Olduvai (Tanzània). 1,8 milions d’anys. Museu Nacional de Dar Es Salaam (Tanzània)
Richard Leakey va descobrir, el 1972, les restes d’un crani (de fet, centenars de trossets d’un crani trobats per B. Ngeneo i reconstruït per M. Epps i A. Walker) a la riba oriental del llac Turkana (Kenya), conegut com el KNM-ER 1470 (Kenya National Museum – East Rudolf 1470); d’una antiguitat d’uns 2 milions d’anys i un volum cranial de 775 cc, pesava uns 40 Kgr i menjava més carn que l’australopitec. Alguns científics afirmen que les diferències de dimensió i de forma indiquen que pertany a una nova espècie, Homo rudolfensis, però aquest té un cervell lleugerament més gran, l’esquelet facial és més gran, ample i pla, el torus està molt poc marcat i les mandíbules i les dents són més grans que les d’Homo habilis. Alguns pensen que KNM-ER 1470 s’hauria d’anomenar Kenyanthropus rudolfensis.
KNM-ER 1470. http://humanorigins.si.edu/evidence/human-fossils/fossils/knm-er-1470
La forma corporal d’Homo habilis la coneixem gràcies a la descoberta, el 1986, per Donald Johanson i Tim White, a Olduvai, d’un esquelet molt fragmentari, l’OH 62.
OH 62. http://truthopia.wordpress.com/2009/03/01/i-science-h-origin-of-man-evolution-style-home-habilis/
L’homo habilis quasi no es distingeix físicament de l’australopitec. És, tanmateix, la més antiga espècie del gènere Homo, l’hàbitat del qual es situa en el continent Africà, i va viure entre els 2.200.000 i els 1.600.000 anys ane, convivint amb l’australopitec en el temps, però no en l’espai. Sembla que fou substituït per l’Homo Erectus.
Característiques
¿Quina és la morfologia bàsica de l’homo habilis?
Entre els 2.200.000 i els 1.600.000 anys s’han trobat a l’Àfrica oriental i a Sud-àfrica fòssils de morfologia més moderna que l’australopitec, i se’ls ha considerat d’una nova espècie: la de l’Homo habilis.
Aquestes són les seves característiques principals:
– crani més arrodonit
– desenvolupament del volum endocranià (entre 500 i 700 cc)
– manca de cresta sagital
– arcada supraorbital reduïda
– rostre poc projectat cap endavant
– dents anteriors més fortes que les laterals i, alhora, més reduïdes que les dels australopitècids: incisives espadiformes i manca de diastema
– foramen magnum ubicat més cap el centre
– dits corbs de peus i mans, el que indica que encara utilitzava els arbres
– talla sistemàtica de la pedra: els còdols amb un tall afilat en una o dues cares són els típics de la cultura olduvaiana o del Paleolític inferior arcaic
– organització de l’espai amb existència de zones d’esquarterament d’animals i zones de talla de pedra
– dieta variada i oportunista, fruit de l’acció de carronyejar i no de la caça organitzada
– la posició bípeda de la femella provoca una reducció de la pelvis que té com a conseqüència un avançament dels parts (cosa que implica prematuració del neonat, un major temps dedicat a la criança i la necessitat de mantenir vincles socials forts que coadjuven al desenvolupament de la cultura)
-el cap del fèmur és més gran, curt i arrodonit
– altura d’uns 1,3 metres i el seu pes uns 40 kgr
Què separa, doncs, l’homo habilis de l’australopitec?
Si l’australopitec va desaparèixer, què va permetre sobreviure a l’homo habilis?
Què és el que ens fa humans?
Troballes realitzades al nord-est Africà (zona del llac Turkana) per Louise i Meave Leakey publicades el 2007 aproximen l’existència de l’Homo habilis fins 1.440.000 anys abans del present. Tal datació implica que almenys durant uns 500.000 anys van coexistir en els mateixos territoris Homo habilis i Homo erectus. Les investigadores opinen que inicialment no tenien per què existir majors conflictes entre les dues espècies, tanmateix el creixement de la població d’Homo erectus hauria acabat amb una lluita pels recursos de la qual haurien sortit reeixits els Homo erectus. D’altra banda, aquesta mateixa troballa posa en dubte una filiació directa entre ambdues espècies, encara que hi ha autors com Erik Trinkaus opinen que la convivència no descarta que Homo habilis sigui avantpassat directe d’Homo erectus.
L’increment cerebral
¿A què pot ser degut el creixement del cervell?
Una diferència fonamental de l’homo respecte l’australopitec és el volum cranial. Quines hipòtesis podem formular per explicar aquest increment cranial?
El creixement del cervell és el resultat de tot un seguit d’activitats interrelacionades, una de les quals, potser la més important, fou la de compartir el menjar. El fet de posar en comú el menjar guarda relació amb l’existència d’una estructura de grup; i fou aquesta cohesió social la que, probablement, va donar avantatges a l’homo per damunt de l’australopitec.
La vida en comú permet desenvolupar noves estratègies per la supervivència, facilita la transmissió d’informacions (tècniques, coneixements sobre plantes i animals…), possibilita la memòria col·lectiva (es poden millorar les capacitats i coneixements existents), i està a la base del desenvolupament del llenguatge. A més, a l’interior del crani del KNM-ER 1470es va descobrir una petita marca allí on els humans actuals hi tenim el centre del llenguatge (el «centre de Broca»); amb això no es vol dir que l’homo habilis ja tingués un llenguatge articulat (això no es donarà fins l’homo sapiens), sinó que segurament ja podia comunicar-se amb alguna cosa diferent als grunys animals. Aquesta cohesió social, generadora de tradicions culturals, incidiria en l’evolució biològica i faria incrementar la capacitat del cervell.
Però, què compartien, bàsicament, els nostres primers avantpassats?
- Per la dentició sabem que eren omnívors; pels estris (destrals de mà i rascadors) sabem que se’n aprofitaven dels cadàvers dels animals grans. L’aprofitament dels cadàvers és una estratègia alimentària en la que hi participen diferents espècies animals sense entrar en conflicte entre elles (les aus de poderosos pics ganxuts obren la pell, les de coll llarg i pic més dèbil prossegueixen el treball…); a més l’aprofitament del cadàver eximeix del gran esforç que comporta l’atac i persecució dels animals vius.
- Per tant, la postura erecta li donava, sobretot, capacitat per a córrer, i la visió aguda li permetia tant el càlcul de distàncies amb precisió com l’observació del vol dels voltors; per altra banda, les eines lítiques, bàsicament, ascles i rascadors, li servien per a esquarterar cadàvers i tallar la carn a tires. Cal tenir present que per al millor aprofitament dels animals morts cal arribar-hi abans que els competidors i en un moment comprès entre la mort de l’animal i l’inici de la seva descomposició.
I què n’obtenia l’homo habilis de l’aprofitament dels cadàvers dels animals? Per una banda, la medul·la i els ossos (els còdols servien, sobretot, per a trencar els ossos i extreure’n la medul·la), i per l’altra, carn. En definitiva, una alimentació rica en fòsfor, element que està a la base de l’augment del cervell.
No podem fer créixer el cervell anant a un gimnàs i desenvolupant-lo com qui fa culturisme. La capacitat del cervell ja està determinada en la formació del fetus; i una base indispensable pel funcionament del cervell és el fòsfor. Si l’alimentació es basa en fulles i fruits, amb un contingut baix en proteïnes i pobre en fòsfor, el desenvolupament del cervell quedarà restringit.
La diferència de la mida cerebral relativa entre els pongins i els hominins significa, segurament, que aquests darrers tenien una font més rica de fòsfor, ja que, contràriament, el seu cervell no hagués pogut créixer. Ara ja sabem d’on obté el fòsfor l’homo habilis: de l’aprofitament dels ossos i de la medul·la dels cadàvers animals [menjar-se el cervell de la presa no té res a veure amb la brutalitat o rituals cranials, sinó en el fet que aquest òrgan és la millor font de compostos de fòsfor].
Per això és tan important l’alimentació que la mare proporciona al fetus: durant l’embaràs i, després, fins al deslletament (3/4 anys) cal proporcionar a la cria agents nutrients de gran valor; i l’embaràs i la producció de llet materna «costa» a l’organisme matern substàncies i energia. La dona necessita, doncs, una alimentació més rica en fòsfor i proteïnes que la de l’home. A l’home només li cal l’alimentació precisa per al seu creixement i una aportació de greixos i hidrats de carboni per dur a terme la tasca de proveïment de carn i vegetals (potser és aquest tipus d’alimentació el responsable que l’home sigui una mica més gran i forçut que la dona). Gràcies al treball cooperatiu de dones i homes es podien satisfer les mútues necessitats alimentàries. En els pobles caçadors-recol·lectors actuals les dones aporten, amb plantes, fruits, insectes i ous, el 75 % de la dieta; la resta, amb carn, l’aporta l’home.
Ara bé, fins on pot créixer el cervell?
Tant com ho permeti l’obertura de la pelvis. La gran mida del cap de la cria obre el canal del part fins els límits del que és possible i suportable. Els dolors de part és el preu que paga la dona pel creixement de la capacitat craniana. Per tal que el crani del nou nat passi a través del canal del part, les cries humanes neixen amb obertures en les plaques òssies (les sutures cranials, fontanel·les), les quals permeten que el cap del nou nat canviï de forma (fins i tot els ossos del crani es poden sobreposar). Aquestes obertures o espais també permeten que el cervell creixi dins dels ossos del crani i s’aniran tancant a mesura que el cervell assoleixi la seva mida completa.
A més de l’increment cerebral, quines interrelacions amb el medi es varen produir per tal de garantir la supervivència de l’homo habilis?
Medi i estratègies de supervivència
¿Quines adaptacions mediambientals ajudaren a sobreviure a l’homo habilis?
Per aprofitar els cadàvers morts, què necessitava l’homo habilis?
Ja hem vist que li calia la capacitat de córrer durant molt de temps per tal d’arribar aviat a la presa. I tots sabem que quan correm ens acalorem i la pell envermelleix (la sang circula més per la perifèria del cos per tal d’alliberar calor, és a dir, per refrigerar-se). El moviment produeix calor i aquesta calor excessiva s’ha d’eliminar si no volem que la temperatura del nostre cos es desestabilitzi. L’ésser humà aconsegueix eliminar l’excés de calor gràcies a la gran superfície del seu cos i al fet que els músculs motors, el panxell i les natges, estan lluny dels òrgans interns.
Però, com l’ésser humà aconsegueix refrigerar el cos de manera suficient mentre corre?
Amb el desenvolupament de les glàndules sudorípares i la pèrdua de pèl corporal. La nostra nuesa corporal respon a la necessitat de refredar el cos: l’aigua extreu calor del entorn quan s’evapora; per això quan més calor tenim més suem. I per això necessitem beure: som el mamífer que més set té (per això depenem de la disponibilitat d’aigua). A la sabana de l’Àfrica tropical l’època de pluges té abundoses precipitacions, a l’època seca els rius porten aigua, i les fonts estan a distàncies raonables.
Però, amb l’evaporació perdem sal (la suor és salada). Quant més suem, més sal consumim. El desenvolupament de les glàndules sudorípares només fou possible en un entorn natural que oferís la suficient sal per compensar la que es perdia. A l’Àfrica oriental la radiació solar és intensa i evapora l’aigua de llacs superficials alliberant sal; a més, a la zona volcànica del Rift neixen aigües superficials riques en sal.
Donada la nuesa corporal, la pell va haver de produir un pigment fosc, la melanina, per protegir-se dels raigs ultraviolats del sol. L’Homo habilis era, doncs, de pell fosca. Els nostres Adams i Eves eren, doncs, negres.
També per aprofitar els cadàvers fan falta estris adequats. I, on es troben les pedres més adients per fabricar-los?
El sílex (o pedra foguera) i l’obsidiana són les millors, i es troben en el curs superior i mitjà de rius de corrent ràpid amb llit de còdols i a les zones volcàniques. L’Homo habilis, doncs, només podia viure en aquells indrets on la naturalesa li proporcionés pedres amb caires afilats. I les zones volcàniques de l’Àfrica oriental n’estan plenes.
Si estava tan ben adaptat, per què va desaparèixer? Cap a on va evolucionar?
Homo rudolfensis
Homo rudolfensis és una espècie d’hominí fòssil, proposada per Valerii Alexeev el 1986, l’espècimen tipus del qual és el KNM-ER 1470, trobat a Koobi Fora (vora oriental del llac Turkana), per Bernard Ngeneo, un membre de l’equip de Richard Leakey, el 1972. Alexeev el va designar el 1986 com Pithecanthropus rudolfensis, encara que els qui el consideren una espècie diferent l’assignen al genero Homo o al gènere Australopithecus.
Alguns paleoantropòlegs dubten que sigui una espècie diferent d’Homo habilis, però no és aquesta l’opinió dominant en l’actualitat, a causa d’unes marcades diferències morfològiques entre ambdós tipus, entre les quals cal esmentar les següents:
- forma de la cara, principalment en la regió supraorbital i malar (de la galta), que presenta molt llarga, profunda i inclinada cap endavant.
- mesures cranials en el seu conjunt: un 45% de les mesures que es van comparar entre les dues espècies superen el dimorfisme sexual dels goril·les.
- volum cranial: al voltant dels 750 cm3, davant els 650 cm3 de l’Homo habilis.
- dentadura: unes dents postcanines més àmplies, amb arrels i corones més complexes i esmalt més gruixut.
L’Homo rudolfensis va habitar en un període de temps comprès entre 2,4 (edat de l’espècimen fòssil UR 501) i 1,9 milions d’anys a l’est de l’Àfrica, per la qual cosa es considera contemporani de l’Homo habilis, així com d’altres hominins, com el Paranthropus boisei. La seva àrea de dispersió sembla haver estat exclusivament l’est Africà, per la qual cosa molts paleoantropòlegs el consideren un endemisme.
A partir del descobriment del Kenyanthropus platyops, s’accepta que Homo rudolfensispodria haver evolucionat a partir de Kenyanthropus, si bé existeixen altres hipòtesis sobre les seves relacions filogenètiques.
Finalment, el fet de la coexistència d’aquestes dues espècies africanes (Homo rudolfensis i Homo habilis), ha portat a alguns autors a suggerir un interessant debat al voltant de la coexistència de dues espècies intel·ligents, ja que sembla ser clara l’associació d’Homohabilis amb les indústries lítiques d’Olduvai i Homo rudolfensis amb les del llac Turkana, que encara no ha arribat a les seves últimes conseqüències.
La troballa de dos fòssils, un os de mandíbula d’Homo habilis d’1,44 milions d’anys i un crani d’Homo erectus de fa 1,55 milions d’anys, al nord de Kenya, per un equip d’experts liderat per Meave i Louise Leakey, a més de demostrar que ambdues espècies van coexistir, ha donat nous arguments per mantenir la hipòtesi segons la qual Homo erectusdescendeix d’Homo rudolfensis. Alguns experts mantenen que Homo erectus i Homorudolfensis van ser espècies germanes descendents d’alguna altra que va viure a Àfrica, de 2 a 3 milions d’anys abans del present.
Homo naledi
El jaciment on s’ha trobat Homo naledi (del sesotho naledi, «estrella») va ser descobert pels espeleòlegs aficionats Rick Hunter i Steven Tucker el 2013 a la cova Rising Star a Sud-àfrica, a uns 800 m del jaciment de Swartkrans. La cambra situada al final de la cova era accessible només a través de dos passos estrets amb menys de vint-i-cinc centímetres d’amplada. Per arribar fins a la cambra cal recórrer uns 80 metres de cova, enfilar una paret i lliscar per una esquerda que els investigadors comparen amb la boca d’una bústia. El descobriment fou anunciat el setembre del 2015 per l’equip de Lee Rogers Berger.
La troballa a la cambra Dinaledi consistia en 15 individus diferents, que comprenen 1.550 exemplars d’ossos. La seva distribució en capes dels ossos suggereix que haurien estat dipositats durant un llarg període de temps, potser fins i tot centúries. Els fòssils inclouen cranis, mandíbules, costelles, dents, els ossos d’un peu gairebé complet, d’una ma, i de una oïda interna. Es va trobar els ossos d’ancians, joves i nens petits. Alguns dels ossos s’assemblen als dels humans moderns, mentre que altres ossos són més primitius que el de l’australopitec (El polze, els ossos del canell i el palmell són similars als modernes mentre que els dits es corben, més com els de l’australopitec, i més útils per a trepar).
Els fòssils encara no s’han datat però es considera que tenen uns 2,5 milions d’anys d’antiguitat. La posició filogenètica de l’Homo naledi és incerta. Tanmateix les característiques anatòmiques el situen proper als primers Homo. Els autors de la descoberta afirmen que es tracta d’una nova espècie d’humà primitiu, però encara no es pot confirmar.
L’espècie ha estat descrita el 2015 por Berger i col·laboradors. Es caracteritza per una massa corporal i una estatura similar a la de les poblacions humanes de cos petit (estatura mitjana de 1,50 m i pes de 45 kgr), però amb un volum cranial (entre 465 cm3 i 560 cm3) semblant al dels australopitecs, i amb una morfologia cranial semblant als exemplars més antics del gènere Homo. L’esquelet combina característiques primitives ja presents en els australopitecs amb característiques derivades presents en els primers homínids. La dentició és primitiva i petita. Les mans tenen adaptacions manipuladores humanes. Així mateix són humanes les característiques de cames i peus. No obstant això, el tronc i extrem proximal del fèmur mostren característiques que l’apropen més als australopitecs.
Cap dels ossos té marques de traumatisme per una possible caiguda a la fossa, ni tampoc signes d’haver estat devorats per un animal o per la seva pròpia espècie, com sí s’esdevé en l’únic jaciment comparable: la Sima de los Huesos a Atapuerca (Burgos).
Controvèrsies científiques
Christoph Zollikofer, de la universitat de Zuric i Tim White, de Berkeley no consideren que es pugui atribuir a una nova espècie, sinó que es podria tractar d’una forma primitiva d’Homo erectus. Per al paleoantropòleg Yves Coppens no és un Homo sinó un Australopitec. Ian Tattersall, del Museu Americà d’Història Natural pensa, també, que els fòssils presenten característiques que els associen més aviat als australopitecs que al gènere Homo. Per a l’antropòleg Jeffrey Schwartz de la universitat de Pittsburgh, els fòssils pertanyen probablement a dues espècies diferents i qüestiona el fet que es tracti d’un ritual funerari. Per la seva banda, Laurent Bruxelles, geòleg, planteja dubtes respecte a fer passar els cossos a través de l’accés actual i considera que hi hauria una altra entrada antigament.
Berger, Lee R (i altres): «Homo naledi, a new species of the genus Homo from the Dinaledi Chamber, South Africa», eLife, 2015;4:e09560, 10 setembre 2015, http://elifesciences.org/content/4/e09560.full
Sample, Ian. «Homo naledi: New species of ancient human discovered, claim scientists». The Guardian, 10 setembre 2015 http://www.theguardian.com/science/2015/sep/10/new-species-of-ancient-human-discovered-claim-scientists
Hervé Morin: «Homo naledi», une découverte qui laisse perplexe
http://www.lemonde.fr/paleontologie/article/2015/09/10/homo-neladi-un-mysterieux-cousin-d-afrique-du-sud_4751937_1650762.html#A0LqwFGGoAJatMzS.99
Le Monde, 10.09.2015, http://www.lemonde.fr/paleontologie/article/2015/09/10/homo-neladi-un-mysterieux-cousin-d-afrique-du-sud_4751937_1650762.html
[1] James Dudley: «The Paleolithic of Hadar, Ethiopia». Stanford University. 1 Juny 2010. En línia a http://jamestdudley.weebly.com/uploads/3/7/9/7/37977239/paleolithic_of_hadar.pdf
[2] Kimbel W.H., Johanson, D.C. i Rak, Y.: «Systematic assessment of a maxilla of Homo from hadar, Ethiopia». Am. H. Phys. Anthropol. 1997 juny; 103 (2), pàgs. 235-62
[3] Kimbel WH. i altres: «Late Pliocene Homo and Oldowan Tools from the Hadar Formation (Kada Hadar Member), Ethiopia». Journal of Human Evolution (1996) 31, 549–561. En línia a http://archaeology.huji.ac.il/depart/prehistoric/erellah/1996_Kimbel_et_al.pdf
[4] L. S.B. Leakey, P.V. Tobias i J.R. Napier: «A new species of the genus Homo from Olduvai Gorge». Nature, Vol. 202, nº 4927, 4 d’abril del 1964, pàgs. 7-9. En línia a http://www.clas.ufl.edu/users/krigbaum/proseminar/leakey_etal_nature_1964.pdf
Joan Campàs Aura digital
Curs: Orígens de l’Art i evolució humana: l’homo significans
Estudis d’Arts i Humanitats de la UOC
Materials del curs en format .pdf
1. Una visió de conjunt de l’art paleolític
2. El paleolític: cronologia i referències etnoarqueològiques
3. Evolució tecnològica en el Paleolític. Referències etnoarqueològiques de l’hàbitat
4. La construcció del coneixement de l’art del Paleolític
5. El paradigma d’Henri Breuil i la seva crisi
6. L’estructuralisme de Leroi-Gourhan i Annette Laming-Emperaire
7. Del xamanisme a l’art com expressió de l’organització social
8. El procés evolutiu: Darwin i la selecció natural
9. Darwin i la selecció sexual. El cicle menstrual
10. La selecció sexual: la competència espermàtica
11. La selecció sexual: l’orgasme femení
12. Sobre els orígens de l’art: les coalicions de dones pintades
13. Una perspectiva ecològica de l’evolució humana
14. Els primers hominoides
15. Els homínids: els preaustralopitecins
16. Els hominins: els australopitecins
17. Paranthropus
18. Els primers Homo: Homo habilis
19. El poblament d’Euràsia: l’homo ergaster/erectus
20. Homo antecessor, rhodesiensis, heidelbergensis i floresiensis
21. L’Homo neanderthalensis
22. L’Homo sapiens
23. Sobre el concepte d’art
24. Les primeres expressions artístiques
25. Sobre els orígens de l’art del Plistocè
26. L’art rupestre del Plistocè
27. Temes i distribució de l’art parietal
28. L’art moble del Plistocè
29. Temes de l’art mobiliari
30. Art mobiliari del Paleolític Superior. Interpretacions
31. Sobre els orígens del llenguatge