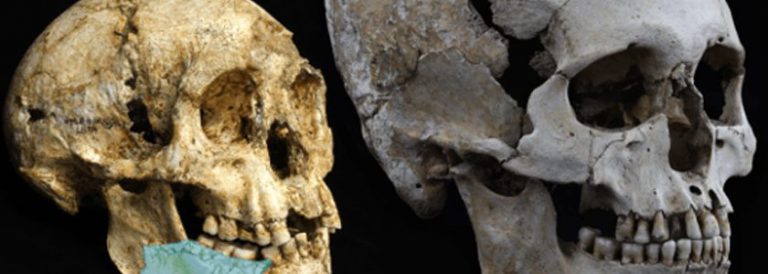
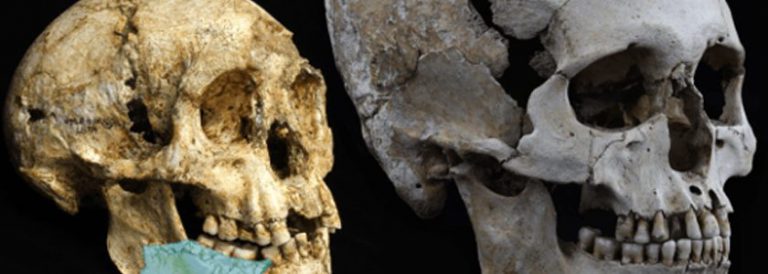
Homo antecessor
Homo antecessor és una espècie fòssil pertanyent al gènere Homo, considerada l’espècie hominina més antiga d’Europa amb una antiguitat de més d’un milió d’anys (Plistocè Inferior). Eren individus alts, forts i amb una cara de trets moderns, encara que el seu cervell fos més petit que el de l’home actual.
El nom d’homo antecessor es va escollir perquè en llatí arcaic «antecessor» significa pioner, i els humans d’Atapuerca són els europeus més antics. En llatí més modern «antecessor» admet l’accepció de predecessor, i la nova espècie és probablement un avantpassat directe de l’Homo sapiens. En assignar aquest nom els autors van voler emfatitzar que els fòssils d’aquesta espècie pertanyen a la primera població d’hominins que va arribar al continent europeu.
La definició d’aquesta espècie és fruit de les més de vuitanta restes trobades des de 1994 en el nivell TD6 del jaciment de Gran Dolina[1] a la Serra d’Atapuerca[2], Espanya. D’acord amb els seus descobridors, entre els caràcters anatòmics d’aquests hominins es pot destacar un conjunt de trets molt primitius en l’aparell dental, que van portar a establir una relació entre aquests i els hominins africans del Plistocè Inferior. Una mandíbula molt ben conservada d’una femella Homo antecessor, d’entre 15 i 16 anys, recuperada del jaciment de la Gran Dolina té similituds molt clares amb les de l’Home de Pequín (Homoerectus), la qual cosa suggereix un origen asiàtic d’Homo antecessor. Tanmateix, el patró de desenvolupament i erupció de les dents és pràcticament idèntic al de les poblacions modernes.
La morfologia facial és similar a la d’Homo sapiens, amb orientació coronal i lleugera inclinació cap enrere de la placa infraorbital que determina la presència d’una fossa canina molt conspícua. La vora inferior d’aquesta placa és horitzontal i lleugerament arquejada. L’arc superciliar és en doble arc i la capacitat encefàlica, estimada a partir d’un fragment incomplet d’os frontal, indica una xifra superior als 1.000 cm3. La morfologia de la mandíbula recorda a la de certs hominins molt posteriors, del Plistocè Mitjà, de l’espècie Homo heidelbergensis, com els de la Sima de los Huesos, també a Atapuerca. L’esquelet postcranial indica una certa gracilitat en comparació amb la major robustesa de l’Homoneanderthalensis, de la segona meitat del Plistocè Mitjà.
A l’actualitat, la validesa d’aquesta denominació com a espècie diferent és defensada pels seus descobridors[3] i altres experts, que consideren que Homo antecessor precedeix a Homo heidelbergensis i per tant és també avantpassat d’Homo neanderthalensis. Tanmateix, part de la comunitat científica la considera una simple denominació, no específica, per a referir-se a restes trobades a Atapuerca, que ells assignen a l’espècie Homo heidelbergensis o bé la consideren una varietat d’Homo erectus/Homo ergaster.
El març de 2008 es van donar a conèixer noves restes d’Homo antecessor, concretament part d’una mandíbula d’un individu d’uns 20 anys i 32 eines de sílex de tipus Olduvaià, datats en 1,2 milions d’anys d’antiguitat, el que fa retrocedir considerablement la presència d’hominins a Europa. Les restes van ser trobades el 2007 a la Sima del Elefante, jaciment situat a uns 200 metres de la Gran Dolina.
De les seves característiques cal esmentar:
- el cervell supera els 1.000 cm3 de capacitat
- més alt i robust que els humans actuals
- nas prominent, mancat de barbeta i front fugisser
- incipient arc supraorbital
- tecnologia similar a l’olduvaiana
Pel que fa a les possibles repercussions en la història evolutiva podem citar, de moment, les següents:
- relega l’homo erectus, considerat per alguns com avantpassat directe de l’homo sapiens, a una via que es va extingir sense descendència en l’arbre evolutiu
- indica que Europa tenia una abundant població humana fa més de 500.000 anys
- pot ser l’avantpassat comú de l’homo neanderthalensis i de l’homo sapiens
Homo cepranensis
L’Home de Ceprano és un exemplar fòssil del crani d’un hominí, descobert per Italo Bidittu el 1994, a la localitat de Ceprano, província de Frosinosne, Itàlia. Després de la reconstrucció dels fragments trobats, realitzada pel geòleg Aldo Segre i la paleontòloga Eugenia Segre Naldini, es calcula que la capacitat craniana d’aquest espècimen podria ser de fins a 1.200 cm3, el que significa un cervell clarament major que el de l’Homo ergaster i l’Homo erectus. S’ha estimat que aquest fòssil data de fa almenys 800.000 anys. Les diferències trobades amb les espècies conegudes d’Homo han portat a la definició formal d’aquest homininí com a espècie diferent: Homo cepranensis.[4]
Homo rodhesiensis
Homo rhodesiensis és una espècie d’hominí fòssil del gènere Homo, trobat per primera vegada el 1921 a la localitat anomenada pels anglesos Broken Hill, actualment Kabwe, a Zàmbia (antiga Rhodèsia del Nord, i per això es va denominar Home de Rhodèsia). Es considera que va viure només a Àfrica, des de fa 600.000 fins 160.000 anys abans del present, durant el Plistocè Mitjà.
La capacitat neurocraneal de l’Homo rhodesiensis era relativament alta, entre 1.280 i 1.325 cm³. Un crani que podria ser el més antic d’aquesta espècie, trobat a Bodo, Etiòpia, datat en 630.000 anys, tenia una capacitat endocraniana de 1.250 cm3. Altres fòssils amb característiques morfològicament similars han estat trobats a Sud-àfrica (Home de Saldaña), Tanzània, el Marroc i Algèria.
Morfològicament conserva certs trets antics comuns a l’Homo erectus, Homo ergaster i Homo antecessor, en té altres que han permès relacionar-lo amb l’Homo heidelbergensis i Homo neanderthalensis, però a més presenta trets que apunten cap a l’Homo sapiens. Per aquestes característiques intermèdies ha estat comparat amb l’Home de Petralona.
Homo heidelbergensis
L’Homo heidelbergensis és una espècie extinta del gènere Homo, que va sorgir fa més de 500.000 anys i va perdurar almenys fins fa 250.000 anys (Plistocè Mitjà). Eren individus alts (1,75 metres) i molt forts (arribarien als 100 quilograms), de grans cranis (1.350 cm3) molt aplanats amb relació als humans actuals, amb mandíbules sortints i gran obertura nasal. Se li va donar aquest nom perquè els primers fòssils van ser descoberts a prop de Heidelberg, Alemanya.
És un avantpassat directe de l’Home de Neandertal a Europa. Tot i que és molt similar als Homo sapiens arcaics trobats a Àfrica (per exemple: Homo rhodesiensis i Homo sapiens idaltu), se sap avui que Homo heidelbergensis no va ser avantpassat directe dels humans moderns. Entre Homo antecessor i Homo neanderthalensis va existir aquesta espècie (Homo heidelbergensis). Presenta en general caràcters intermedis entre Homoerectus/Homo ergaster i Homo sapiens.
Presenta un tors occipital dividit (en l’Homo erectus el tors, o cresta, és continu) i una gran capacitat neurocraneal. Si en els Homo erectus la dieta probablement va estar basada en el cleptoparasitisme, el carronyeig (que aportaven proteïnes i greixos de bona qualitat) i un règim d’aliments vegetals, en l’Homo heidelbergensis es fa patent un predomini de la dieta carnívora amb evidències de cacera. La pressió evolutiva perquè entre els heidelbergensises desenvolupés la cacera hauria estat en les condicions ecològiques del territori que van colonitzar: Europa era freda i durant sis mesos existien molts menys recursos alimentaris obtenibles de vegetals que a Àfrica, per això el carronyeig i el cleptoparasitisme (robar preses als animals predadors) no aportava suficients nutrients. Això hauria induït (per selecció) a l’aparició de conductes socials dedicades a la cacera: bandes d’heidelbergensis s’organitzaven per perseguir altres animals i ultimar-los en trampes naturals (precipicis, pantans) o llançant-los grans destrals lítiques bifaços i, fins i tot, rústics venables de fusta afuada.
El més antic dels fòssils de l’espècie és una mandíbula inferior trobada pel treballador d’una mina a Mauer, a prop d’Heidelberg. Posteriorment, en una cova anomenada Cauna de l’Aragó, a França, es van trobar les restes fragmentàries d’una dotzena d’individus. Un dels més complets és la cara i par de la caixa craniana d’un individu conegut com a Home de Talteüll, que té una gran semblança amb el crani de l’Home de Petralona, trobat en una cova a Grècia. Altres llocs on s’han trobat fòssils d’aquesta espècie són Steinheim (Alemanya), Swanscombe (Anglaterra) i la Sima de los Huesos a la Serra d’Atapuerca (Espanya), on es van trobar 5.000 fòssils pertanyents a un mínim de 32 individus, que daten de fa 400.000 anys considerats avantpassats dels neandertals, restes molt bé conservades; entre ells destaquen el crani número 5 (anomenat popularment Miguelón) complet, i una pelvis molt bé conservada d’un individu conegut popularment com Elvis. A la Xina s’han trobat fòssils que concorden amb aquest grup, a Dali: un crani de fa 280.000 anys, i un esquelet a Jinniushan.
A subratllar que a Terra Amata (França), es van trobar restes de refugis construïts durant l’època d’Homo heidelbergensis. Aquests refugis tenien una forma ovalada, d’uns vuit metres de llarg i set d’ample. Dins del refugi s’hi va trobar restes de cendra.
S’han trobat fòssils relacionats amb aquesta espècie a l’Àfrica a Bodo, al nord-est d’Etiòpia, amb un exemplar més robust però similar que data de fa 600.000 anys, i a Kabwe, Zàmbia. També al llac Ndutu, Tanzània. Els ossos postcranials concorden amb els d’Aragó, i indiquen que aquesta era una forma robusta, però moderna. Els científics estan dividits en la seva interpretació sobre aquests fòssils africans. Alguns els situen en l’espècie Homo heidelbergensis. D’altres pensen que els fòssils europeus i africans pertanyen a dues espècies diferents, i només les poblacions africanes, classificades com a Homo rhodesiensis o Homo sapiens arcaic, van evolucionar fins a l’Homo sapiens. Tampoc no hi ha unanimitat sobre la interpretació de l’Home de Petralona (Grècia), que podria ser un Homo heidelbergensis tardà o més bé un Homo rhodesiensis o un altre Homo sapiens arcaic que va arribar a Europa des d’Àfrica, per les costes de la Mediterrània.
Fa aproximadament mig milió d’anys es van començar a gestar els trets morfològics que en uns centenars de milers d’anys van donar origen al neandertal. Els preneandertales o Homo heidelbergensis mostraven una creixent corpulència física, expressada en l’augment de la talla, 1,75 a 1,80 metres i prop de 100 quilograms de pes (mitjana masculina). Les mandíbules també tenien una gran força i robustesa a causa de l’estrès al qual eren sotmeses.
El conjunt d’aquests fòssils revelen un hominí amb un crani més «inflat» que el de cap Homo erectus/Homo ergaster, encara que posseeix arcs supraciliars. La part posterior del crani és més arrodonida que en l’Homo erectus/Homo ergaster, i les galtes són inflades, com en els neandertals, encara que la cara és més plana. La seva capacitat neurocraneal no dista gaire de la de l’home modern, sobre els 1.350 cm3. Una altra semblança amb l’Homo sapiens actual era el seu aparell fonador. Això va fer pensar que el llenguatge entès d’una manera potser diferent de l’actual, ja era present en aquests grups.
Els estris associats amb els fòssils consisteixen bàsicament en «pedres de tallar» i algunes eines d’ascles com puntes i raspadors de fusta, os i banya, sent els iniciadors d’aquesta tècnica. Dins de la seva tecnologia trobem el bifaç, també anomenat destral de mà; és de grans dimensions i està tallat per ambdues cares. La seva utilitat és molt diversa. Segons l’anàlisi d’altres pedres mostren un ús per adobar pells o treballar la fusta. Fa 400.000 anys ja usaven rudimentàries llances de fusta. També és molt probable que en aquesta època el foc fos utilitzat i gradualment fos generalitzat, ja que hi ha evidències de fogueres.
Dins d’aquesta espècie també podem començar a parlar de petits grups amb una cohesió social, ja que segons estudis paleontològics de diferents restes s’han trobat, per exemple, mandíbules totalment edentades; això indica que l’ancià, després de perdre la seva dentadura, va continuar vivint ajudat pels seus congèneres. Això es tradueix en una conducta diferencial a l’hora del tractament dels seus cadàvers (Sima de los Huesos). L’estudi de les dentadures dels heidelbergensis també revela un altre aspecte evolutiu de gran interès: els molars i la dentadura definitiva apareixien entre els 5 i 6 anys després de néixer (en un ximpanzé actual això s’esdevé cap als 3 anys). Si es considera que un individu no pot alimentar-se pel seu compte fins a tenir ben desenvolupada la seva dentició, això indueix a pensar que els heidelbergensis estaven caracteritzats per una prematuració que implicava una prolongada cura de les cries, amb el possible establiment de fortes relacions afectives entre els individus.
L’existència d’aquestes fortes relacions afectives hauria cohesionat les societats heidelbergensis d’una manera que els capacitava per a, per exemple, desenvolupar eficaces estratègies de cacera i de supervivència en general.
Homo floresiensis
Homo floresiensis[5] (Home de Flores) és el nom proposat per a una possible espècie recentment descrita del gènere Homo, extraordinària per la petita mida del seu cos i el seu cervell, i per la seva recent supervivència, doncs es creu que va ser contemporània amb els humans moderns a l’illa indonèsia de Flores. Es va descobrir un esquelet subfòssil, datat fa 18.000 anys, molt complet excepte pels ossos del braç, que encara no s’havien trobat, en jaciments a la cova de Ling Bua el 2003. Posteriorment, al mateix lloc es van recuperar parts d’uns altres sis individus, tots diminuts, així com els ossos del braç dret pertanyents a l’exemplar original i eines de pedra igualment petites d’estrats compresos d’entre 90.000 a 13.000 anys d’antiguitat.
Arbre filogenètic amb les possibles escenaris evolutius; il·lustra alguns dels possibles ancestres de l’Homo floresiensis: Homo erectus asiàtic o Homo erectus d’Àfrica[6].
L’illa de Flores ha estat descrita com «una mena de món perdut», on animals arcaics, llargament extints a la resta del món, havien evolucionat a formes gegants i nanes per especiació al·lopàtrica[7]. L’illa tenia elefants nans (una espècie del gènere Stegodon) i llangardaixos gegants similars al drac de Komodo, així com l’Homo floresiensis, que pot ser considerat una espècie d’humà nan.
Aquest descobriment ha estat objecte d’una gran controvèrsia al món científic, ja que no tots els investigadors estan a favor del descobriment d’una nova espècie, i s’han publicat investigacions a favor i en contra de l’Homo floresiensis[8].
Els primers (i per ara únics) espècimens van ser descoberts per un equip conjunt de paleoantropòlegs australians i indonesis que buscava a l’illa de Flores evidències de la migració humana original d’Homo sapiens des d’Àsia cap a Austràlia. No esperaven trobar una nova espècie, i van quedar bastant sorpresos en recuperar les restes pel cap baix de set individus no Homo sapiens, d’entre 38.000 a 13.000 anys d’antiguitat, de la cova Liang Bua a Flores. Un os del braç, provisionalment assignat a l’Homo floresiensis, té uns 74.000 anys d’antiguitat. També són àmpliament presents en aquesta cova sofisticats estris de pedra d’una mida considerada adient per a un humà d’un metre d’estatura. Els esmentats estris semblen tenir una antiguitat entre 95.000 i 13.000 anys i estan associats amb estegodonts joves (un tipus extingit d’elefant), presumiblement preses de l’Home de Flores.
Els espècimens no estan fossilitzats, sinó que tenen el que ha estat descrit com «la consistència del paper mullat assecant-se » (un cop exposats, els ossos s’han de deixar assecar abans que puguin ser extrets).
Els investigadors esperen trobar ADN mitocondrial en bon estat per comparar-ho amb mostres d’espècimens no fossilitzats d’Homo neanderthalensis i Homo sapiens similars. Tanmateix, la probabilitat que s’hagi conservat ADN és baixa, ja que aquest es degrada més ràpidament en entorns tropicals temperats, on se sap que no es conserva més que algunes dotzenes d’anys. La contaminació de l’entorn proper sembla altament probable donat l’ambient humit en el qual van ser trobats els espècimens.
L’Homo erectus, assenyalat com l’avantpassat immediat de l’Homo floresiensis, tenia aproximadament la mateixa talla que una altra espècie descendent, la dels humans moderns. Tanmateix, els especialistes creuen que a conseqüència del limitat aportament alimentari de l’illa de Flores, l’Homo erectus, arribat a aquest territori fa uns 500.000 anys, va sofrir un fort nanisme illenc (una forma d’adaptació evolutiva també present a l’illa en diverses espècies, incloent el estegodont nan, i observada també en altres illes petites). A part de la diferència de mida, els espècimens semblen semblants en les seves característiques a l’Homo erectus, del que se sap vivia al sud-est asiàtic en la mateixa època que les troballes més antigues de l’Homo floresiensis. Aquestes semblances observades formen la base de l’establiment de la relació filogenètica suggerida.
Tanmateix cal tenir en consideració que no s’han trobat a l’illa restes corresponents a l’Homo erectus, i molt menys formes de transició, trobant-se fins al moment només proves materials (eines de pedra), atribuïbles només presumptivament a una possible ocupació de l’Homo erectus fa 840.000 anys, no descartant-se també que siguin atribuïbles a una altra possible espècie d’hominí existent en aquella època.
El crani LB1 (centre) en comparació amb l’Homo habilis de Kenya (1,9 Ma) (esquerra) i el crani d’Homo erectus de la República de Geòrgia (1,8 Ma) (dreta).
Així, l’espècimen tipus d’aquesta espècie trobada, un esquelet[9] bastant complet, i un crani gairebé complet d’una femella de 30 anys d’edat i prop d’un un metre d’altura, no solament presenta una dràstica reducció en comparació amb l’Homo erectus, sinó fins i tot una talla una mica menor que la de l’Australopithecus, un avantpassat tres milions d’anys més antic i que no es pensava prèviament que s’hagués expandit més enllà d’Àfrica. Això tendeix a qualificar l’Homo floresiensis com el membre més «extrem» de l’extensa família humana, ja que serien certament els més baixos i petits[10].
En relació a l’estatura, l’Homo floresiensis és també bastant diminut comparat amb la mida de l’ésser humà modern. L’altura estimada d’un Homo floresiensis adult és considerablement menor que l’altura mitjana adulta de totes les poblacions humanes modernes físicament més petites, tals com els pigmeus africans (<1,5 metres), Twa, Semang (1,37 metres per a les dones adultes) o els andamanesos (1,37 metres per a les dones adultes). La massa és normalment considerada més important biofísicament que una mesura unidimensional d’altura, i per l’esmentada mesura, a causa dels efectes d’escala, les diferències són fins i tot majors. S’ha estimat que en l’espècimen tipus d’Homo floresiensis era d’uns 25 quilograms.
L’Homo floresiensis a més tenia braços relativament llargs, potser per permetre a aquesta petita criatura enfilar-se cercant la seguretat dels arbres quan ho necessitava. Aquests ossos del braç fan que les inevitables comparacions amb els humans moderns acondroplásics (sobre 1,2 metres) o altres nans no siguin vàlides, ja que aquestes persones no són proporcionalment més petites que les altres per regla general, sinó que només tenen els seus membres més curts.
A més d’una talla corporal petita, l’Homo floresiensis tenia un cervell extraordinàriament petit[11]. L’espècimen tipus, amb 380 cm3, està en el mateix rang que els ximpanzés o els antics Australopithecus. El seu cervell està reduït considerablement respecte al del presumpte antecessor immediat d’aquesta espècie, l’Homo erectus, que amb 980 cm3tenia més del doble de volum cerebral que la seva espècie descendent. Tanmateix, la relació cervell-massa corporal que presenta l’Homo floresiensis és comparable a la de l’Homo erectus, la qual cosa indica que és improbable que les espècies difereixin en intel·ligència. De fet, els descobridors han associat a l’espècie alguns comportaments avançats.
Aquests comportaments estarien associats a l’existència d’evidències de l’ús del foc per cuinar. L’espècie també ha estat relacionada amb eines de pedra de la sofisticada tradició del Paleolític Superior típicament associada amb els humans moderns, que amb 1.310-1.475 cm3 gairebé quadrupliquen el volum cerebral de l’Homo floresiensis (amb una massa corporal incrementada en un factor de 2,6). Algunes d’aquestes eines van ser aparentment usades en la cacera necessàriament cooperativa del Stegodon nan local per aquesta petita espècie humana. Igualment en un altre jaciment anomenat Mata Menge, l’investigador Adam Brumm i els seus col·laboradors han detectat que les eines trobades tenen importants similituds amb les trobades a Liang Bua; presentant les eines de Mata Menge una datació que dóna la sorprenent data d’entre 840.000 i 700.000 anys. Aquestes dates, indiquen que Homo sapiens no va poder fabricar les eines descobertes a Mata Menge, i amb això probablement també les de Liang Bua, ja que l’espècie Homo sapiens no existia en aquella època.
A més, l’illa de Flores va romandre aïllada durant l’edat de gel més recent a causa d’un profund estret, malgrat el baix nivell marí que va unir a bona part de la resta de Sundaland. Aquest fet ha portat els descobridors de l’Homo floresiensis a concloure que l’espècie o els seus antecessors només van poder haver assolit l’illa aïllada per mitjà d’algun transport marítim, potser arribant en rais de bambú fa uns 100.000 anys.
Aquestes evidències observades de tecnologia avançada i cooperació a un nivell humà modern ha impulsat els descobridors a proposar que Homo floresiensis tindria gairebé amb total certesa llenguatge. Aquests suggeriments han resultat ser els més controvertits de les troballes dels descobridors, malgrat la probablement alta intel·ligència de l’Homofloresiensis.[12]
L’altre aspecte notable de la troballa és que es creu que aquesta espècie ha sobreviscut a l’illa de Flores tan recentment com fins fa 12.000 anys. Això la fa l’hominí més longeu, sobrevivint llargament a la desaparició dels neandertals fa uns 28.000 anys. L’Homofloresiensis certament va coexistir amb els humans moderns, que van arribar a la regió fa entre 35.000 i 55.000 anys, durant un llarg període, però es desconeix com van poder haver interactuat.
L’anàlisi de la geologia local suggereix que una erupció volcànica a Flores va ser la responsable de la desaparició de l’Homo floresiensis en la part de l’illa sota estudi fa aproximadament 12.000 anys, junt amb gran part de la fauna local, inclòs el Stegodon nan i les rates gegants de l’illa.
Els descobridors sospiten, tanmateix, que aquesta espècie pot haver sobreviscut molt més temps en altres parts de l’illa de Flores fins a arribar a ser l’origen i font de les històries sobre els Ebu Gogo explicades entre els vilatjans. Es diu que els Ebu Gogo eren cavernícoles petits, peluts i de llenguatge pobre, i presentarien la mida de l’Homofloresiensis. Es creia àmpliament en la seva existència en l’època de l’arribada dels holandesos fa cinc-cents anys, i fins i tot algunes històries diuen que aquestes estranyes criatures han estat vistes fa tan sols un segle.
De forma similar, a l’illa de Sumatra circulen llegendes sobre un humanoide d’un metre d’altura, l’Orang Pendek, encara que són pocs els investigadors professionals que les prenen seriosament, rebent més atenció de l’autodenominada criptozoologia (disciplina l’enfocament de la qual sol ser pseudocientífic). Els investigadors que han treballat en l’home de Flores, han assenyalat que l’Orang Pendek i altres humanoides salvatges del folklore d’Àsia sud-occidental podrien estar relacionats d’alguna forma amb relats sobre antics encontres amb Homo floresiensis o altres hominins nans encara desconeguts.
El descobriment és considerat àmpliament com el més important de la seva classe en la història recent, i va ser tota una sorpresa per a la comunitat antropològica. La nova espècie desafia moltes de les idees de la seva disciplina. Des del segle XIX, quan van començar a descobrir-se els primers neandertals, no s’havien descobert altres hominins coetanis de l’Homo sapiens.
L’Homo floresiensis és tan diferent en forma a altres membres del gènere Homo que obliga al reconeixement d’una possible nova i inimaginable variabilitat en aquest grup, i reafirma una tendència intel·lectual lluny de la idea de l’evolució lineal.
[1] «Los fósiles humanos del nivel td-6 de la Gran Dolina: ‘Homo antecessor’». Pàgina web de Juan Luis Arsuaga y el equipo de investigación Centro UCM-ISCIII. En línia a http://www.atapuerca.tv/atapuerca/fosiles_dolina
[2] J.M. Parés ,L. Arnold, M. Duval, M. Demuro, A. Pérez-González, J.M. Bermúdez de Castro, E. Carbonell, J.L. Arsuaga: « Reassessing the age of Atapuerca-TD6 (Spain): new paleomagnetic results». Journal of Archaeological Science, Volum 40, Issue 12, Desembre del 2013, Pàgs. 4586–4595. En línia a http://www.sciencedirect.com/science/article/pii/S0305440313002197
[3] http://www.diariodeatapuerca.net/Homoantecessor.pdf
[4] Clarke R.J (2000): «A corrected reconstruction and interpretation of the Homo erectus calvaria from Ceprano, Italy», Journal of Human Evolution, Volume 39, Number 4, October 2000, pp. 433-442.
[5] Baab, K. L. (2012) «Homo floresiensis: Making Sense of the Small-Bodied Hominin Fossils from Flores» . Nature Education Knowledge. 3(9):4. En línea a http://www.nature.com/scitable/knowledge/library/homo-floresiensis-making-sense-of-the-small-91387735
[6] Argue, D. et al.: «Homo floresiensis: a cladistics analysis». Journal of Human Evolution57, 623-639 (2009).
[7] Es coneix per especiació alopàtrica (o alopàtrida) a l’especiació per aïllament geogràfic. Del grec állos, diferent, fa referència al mecanisme pel qual una espècie n’origina un altra o altres en àrees diferents.
Aquest fenomen implica la separació d’una població per aïllament geogràfic i la seva permanència de forma aïllada. Amb el temps es produeix una divergència genètica i/o morfològica que deriva en una nova espècie. Un clar exemple és el de les dues espècies de ximpanzés, el ximpanzé comú (Pan troglodytes) i el ximpanzé nan o bonobo (Pan paniscus). Originalment constituïen una sola espècie, però la frontera natural que constitueix el riu Congo va separar les dues poblacions, impedint l’intercanvi de gens entre elles, el que va donar lloc a la especiació observada.
[8] Carlos A. Marmelada (2007). Homo floriesensis. En línia a http://www.educarm.es/templates/portal/images/ficheros/etapasEducativas/secundaria/10/
secciones/425/contenidos/7187/homo_floresiensis.pdf
[9] Larson, S. G. et al :. «Descriptions of the upper limb skeleton of Homo floresiensis». Journal of Human Evolution 57, 555-570 (2009).
[10] Jungers, W. L. et al.: «The foot of Homo floresiensis». Nature 459, 81-84 (2009).
[11] Martin, R. D. et al.: «Comment on “The Brain of LB1, Homo floresiensis”.» Science312, 999 (2006). Veure també Falk, D. et al.: «The brain of LB1, Homo floresiensis». Science 308, 242 – 245 (2005).
[12] Aiello, L.C. «Five years of Homo floresiensis». American Journal of Physical Anthropology 142, 167-179 (2010).
Joan Campàs Aura digital
Curs: Orígens de l’Art i evolució humana: l’homo significans
Estudis d’Arts i Humanitats de la UOC
Materials del curs en format .pdf
1. Una visió de conjunt de l’art paleolític
2. El paleolític: cronologia i referències etnoarqueològiques
3. Evolució tecnològica en el Paleolític. Referències etnoarqueològiques de l’hàbitat
4. La construcció del coneixement de l’art del Paleolític
5. El paradigma d’Henri Breuil i la seva crisi
6. L’estructuralisme de Leroi-Gourhan i Annette Laming-Emperaire
7. Del xamanisme a l’art com expressió de l’organització social
8. El procés evolutiu: Darwin i la selecció natural
9. Darwin i la selecció sexual. El cicle menstrual
10. La selecció sexual: la competència espermàtica
11. La selecció sexual: l’orgasme femení
12. Sobre els orígens de l’art: les coalicions de dones pintades
13. Una perspectiva ecològica de l’evolució humana
14. Els primers hominoides
15. Els homínids: els preaustralopitecins
16. Els hominins: els australopitecins
17. Paranthropus
18. Els primers Homo: Homo habilis
19. El poblament d’Euràsia: l’homo ergaster/erectus
20. Homo antecessor, rhodesiensis, heidelbergensis i floresiensis
21. L’Homo neanderthalensis
22. L’Homo sapiens
23. Sobre el concepte d’art
24. Les primeres expressions artístiques
25. Sobre els orígens de l’art del Plistocè
26. L’art rupestre del Plistocè
27. Temes i distribució de l’art parietal
28. L’art moble del Plistocè
29. Temes de l’art mobiliari
30. Art mobiliari del Paleolític Superior. Interpretacions
31. Sobre els orígens del llenguatge