Evolució del cervell (3) (Som les nostres connexions 13)
14 juliol, 2016
Reflexió inicial
Falta molta informació sobre els primers estadis de l’evolució humana. N’hi ha prou que apareguin uns quants fòssils de certa antiguitat en algun jaciment africà per crear un debat científic sobre la seva possible identitat com a representants de la genealogia humana. Amb independència d’aquestes discussions, els primers quatre milions d’anys de l’evolució humana aparenten tenir poc a oferir pel que fa a modificacions anatòmiques de certa rellevància. Potser sigui per falta d’informació. Encara que la nostra gran explosió evolutiva va passar molt més tard, l’estudi de la diversitat dels hominins del Pliocè és força interessant. Ens interessa esbrinar com va evolucionant el cervell, en quins contextos, per a quines adaptacions i quines funcions pot dur a terme.
Si les relacions filo genètiques de les diferents poblacions del gènere Homo han estat i seguiran sent molt controvertides, tot i disposar d’un registre fòssil relativament abundant, què podem esperar quan amb prou feines comptem amb un grapat de fòssils representant a desenes, si no centenars de milers d’anys?
Primer parlarem dels fòssils africans més importants amb una cronologia d’entre una mica més de 6 i 2 milions d’anys. En aquest rang temporal van viure els primers representants del gènere Homo. En un següent tema ens centrarem en el procés que ens porta de l’Homo habilis al neandertal.
En nom d’una didàctica més senzilla, el discurs que seguirem repassarà de manera breu les troballes no per la data del seu descobriment sinó per la seva antiguitat.
Els primers indicis
Cfr. Tema 15. Els homínids: els preaustralopitecins, del curs en línia Orígens de l’art i evolució humana: l’homo significans. En línia a http://cv.uoc.edu/adf/~04_999_01_u07/llico_15.pdf
Orrorin tugenensis
El 2001 Brigitte Senut [1] i altres investigadors, recolzats per Yves Coppens, van publicar a la revista Comptes Rendus de l’Acadèmia de París la troballa de 14 restes fòssils humans datats en uns 6 milions d’anys; és a dir, la seva cronologia estaria a l’entorn del rang temporal que els genetistes assumeixen per a la separació de la genealogia humana i la dels ximpanzés. Aquests fòssils, doncs, estarien molt propers a l’avantpassat comú d’uns i altres.
Els fòssils descrits per Senut i els seus col·legues van ser trobats en la formació geològica Lukeino, prop de la localitat de Kapsomin, al districte kenyà de Baringo. Els fòssils van ser assignats a un nou gènere i espècie: Orrorin tugenensis [2]. El nom específic indica la procedència dels fòssils, trobats en les turó de Tugen, mentre que el nom genèric significa «Home original» en la llengua de l’ètnia Tugen, que pertany al grup kenià dels Kalenjin.
Els molars d’aquests fòssils són petits respecte a la seva mida corporal i tenen l’esmalt gruixut, en contraposició al que s’observa en els ardipitecs. Aquest fet va portar a Senut i als seus col·laboradors a concloure que els ardipitecs, que posseeixen esmalt fi en els seus molars, s’haurien d’excloure de la genealogia dels hominins i relacionar-se més bé amb la genealogia dels goril·les (tribu Gorillini).
La morfologia del fèmur fa pensar que l’Orrorin tenia locomoció bípeda, però la de l’húmer fa pensar en capacitats. enfiladisses. Les dades disponibles són, tanmateix, escasses com per pronunciar-se sobre el seu possible bipedisme, un tema crucial per als orígens humans.
Sahelanthropus tchadensis
El 2002, Michel Brunet [3] va publicar a la revista Nature la possible evidència del suposat membre més antic de la genealogia humana conegut fins a la data. Aquesta vegada es tractava d’un crani, un parell de fragments de mandíbula i diverses dents. El crani presenta un cert aixafament i distorsió produït pel pes dels sediments.
Cal subratllar la seva localització, la localitat de Toros-Menalla, situada a la regió oest del desert de Djurab, al nord del Txad i a una distància de 2.500 quilòmetres a l’oest de la Gran Vall del Rift. Fins a aquest moment tots els primitius ancestres de la nostra genealogia s’havien trobat a l’est d’Àfrica, on es considera que va viure el nostre avantpassat comú amb els ximpanzés. És evident que a finals del Miocè el clima d’aquesta regió no era el mateix que en l’actualitat, a jutjar per les espècies de mamífers que van viure en aquell lloc fa entre 7 i 6 milions d’anys.
Tenint en compte el lloc de la troballa, l’antiguitat dels fòssils i la combinació peculiar de caràcters, Brunet i els seus col·laboradors van prendre la decisió de nomenar un nou gènere i espècie, Sahelanthropus tchadensis, recordant que el descobriment s’havia realitzat a la regió del Sahel i a la República del Txad. De manera col·loquial es coneix a aquest fòssil com «Toumai», que en l’idioma kanuri de la regió significa «esperança de viure» i s’utilitza per descriure als nens que neixen a l’estació seca.
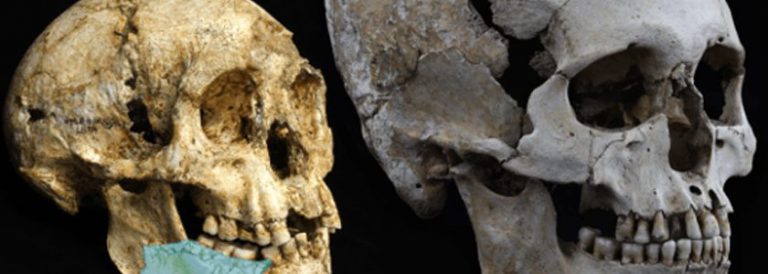
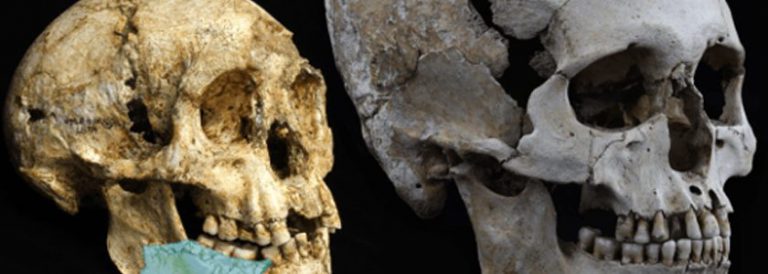
Crani fòssil de 7 milions d’anys, quasi complet, descobert el 19 de juliol del 2001, en el desert al nord del Txad
D’entre els molts caràcters que s’han pogut descriure, en destaquem els més significatius. El crani mostra un tor supraorbitari continu, les seves canines no són massa grans, i no mostra una disposició característica dels simis antropoides (del grec ανθρωποιδης, anthropos, “home” i oides, “similar a”) que rep el nom de «complex C/P3» associada a diastemes al maxil·lar i la mandíbula. El gruix de l’esmalt de les dents és intermedi entre el que s’observa en els hominins i el dels ximpanzés.
El crani de Sahelanthropus va allotjar un cervell d’una mida similar al dels ximpanzés. La capacitat cranial es va poder estimar entre 320 i 380 centímetres cúbics, sense major precisió a causa de la distorsió de l’exemplar. Cal assenyalar la presència d’un foramen magnum (forat magne) més llarg que ample i orientat de manera horitzontal. La seva morfologia i situació a la base del crani és similar a la de hominins del Pliocè i a la del propi gènere Homo, el que suggereix una posició dreta i bipedestació. Per Brunet i els seus col·laboradors no hi ha dubte que ens trobem davant d’un ancestre de la genealogia humana. De confirmar-se, faria endarrerir la divergència evolutiva entre humans i ximpanzès més enllà dels 6 milions d’anys.
Ardipithecus kadabba
Haile-Selassie [4] va trobar les primeres restes d’aquesta espècie al marge oest de la vall del riu Awash. La seva cronologia sembla ben establerta entre 5,8 i 5,2 Ma. La primera decisió d’Haile-Selassie va ser incloure les primeres restes a la subespècie Ardipithecus ramidus kadabba. No obstant això, després de realitzar noves troballes i acarar-se amb els de l’espècie Ardipithecus ramidus, HaileSelassie, Gen Suwa i White van prendre la decisió d’elevar les restes fòssils a la categoria d’espècie. Les diferències temporals i morfològiques entre les dues formes d’ardipitecs semblaven garantir la distinció específica.
En la seva primera descripció d’Ardipithecus kadabba, Haile-Selassie refereix una combinació de caràcters dentals [5] primitius compartits amb primats del Miocè i caràcters derivats, que suggereixen la seva relació amb la genealogia humana. Aquest investigador insisteix que diversos dels caràcters de l’esquelet postcranial dels fragments d’húmer i del cúbit són molt similars als d’Australopithecus afarensis. No obstant això, les evidències del possible bipedisme d’Ardipithecus kadabba només poden estudiar-se en una falange del peu.
Troballes posteriors van donar com a resultat el descobriment de sis noves dents. Dos d’aquests espècimens, trobats a la localitat d’Asa Koma, van revelar una disposició de primer premolar i del caní similar a la dels simis antropoides (complex C/P3). Aquesta va ser la principal raó per distingir a l’espècie Ardipithecus kadabba de la seva possible successora, Ardipithecus ramidus.
Si el complex C/P3 apareix en Ardipithecus kadabba, llavors per què no apareix en hominins anteriors com Orrorin i Sahelanthropus? Haile-Selassie, Suwa i White pensen que aquests dos hominins no pertanyen a la nostra genealogia i podrien representar variacions a nivell específic o subespecífic d’un mateix gènere. Per a aquests investigadors es podria establir una relació filogenètica de la nostra genealogia a Etiòpia, formada pel gènere Ardipithecus, i les espècies Australopithecus anamensis i Australopithecus afarensis, de la qual quedarien exclosos tant Orrorin com Sahelanthropus.
Com bé podem imaginar, el debat entre els científics responsables d’aquestes troballes ha estat tot menys amistós. Aquest debat no fa sinó confirmar que les evidències sobre els primers estadis dels nostres orígens són molt escasses. Tan sols podem afirmar que si els hominins esmentats fins al moment representen veritables ancestres del llinatge humà, hem de reconèixer que la taxa evolutiva durant els dos primers milions d’anys va ser molt lenta, gairebé inapreciable en el registre fòssil.
Ardi i Lucy: ¿mares de la humanitat?
Cfr. Tema 16. Els hominins: els australopitecins del curs en línia Orígens de l’art i evolució humana: l’homo significans. En línia a http://cv.uoc.edu/adf/~04_999_01_u07/llico_16.pdf
Ardipithecus ramidus
La primera publicació sobre les troballes de l’espècie Ardipithecus ramidus és de 1994 (fou descobert el 1992 per Alamayehu Asfaw a Etiòpía), els descobriments més importants van arribar en anys posteriors i es van publicar el 2009, en un número monogràfic de la revista Science, on s’hi descriu la morfologia i la biologia de 35 individus d’aquesta espècie. La majoria de les restes estan constituïdes per fragments de cranis. L’exemplar més complet, a qui van posar el nom d’«Ardi», va pertànyer a una femella l’estatura i pes de la qual es van estimar en uns 120 centímetres i 50 quilograms, respectivament. La pelvis i els peus d’Ardi ens revelen que aquesta espècie ja s’havia posat dreta sobre les extremitats posteriors i que la seva locomoció era bípeda. Molt possiblement els membres d’aquesta espècie no eren capaços de córrer com ho fem nosaltres, però tampoc ho necessitaven en l’hàbitat boscós que van freqüentar.
Aquesta espècie va existir fa 4.4 milions d’anys. Va viure en ambient de bosc tropical plujós temperat, el que indica que si els membres d’aquesta espècie van presentar un bipedisme o semibipedisme, aquest no estava associat encara a la seva expansió a l’hàbitat de sabana.
L’esquelet del gènere Ardipithecus presenta adaptacions tant a la locomoció bípeda com a la possibilitat d’enfilar amb facilitat. La seva pelvis o les extremitats són molt particulars i diferents a les que observem en l’esquelet d’un humà actual. La baixa estatura d’Ardipithecus facilitava les seves possibilitats de moure pels boscos d’Àfrica i d’enfilar-se als arbres davant de qualsevol perill o per trobar menjar.
La mà d’Ardi té dits llargs i un polze no oposable als altres: no estava preparada per a la pinça de precisió, sinó per agafar amb força. Les canines i les premolars encara tenien la morfologia característica dels simis antropoides, però sense diastemes desenvolupats. Aquest aspecte anatòmic i la morfologia de les canines permet pensar en una clara relació filogenètica entre Ardipithecus kadabba i Ardipithecus ramidus.
Australopithecus anamensis
El 1995, un any més tard de la publicació de les primeres restes fòssils d’Ardipithecus ramidus, es va conèixer la troballa de dotze espècimens trobats per l’equip de Meave Leakey a les localitats de Kanapoi i Allia Bay, entre dos dels rius que desemboquen en el llac Turkana. La cronologia d’aquests fòssils és molt fiable, perquè els sediments que els contenen han estat explorats durant anys i les capes d’origen volcànic han estat datades mitjançant el mètode dels isòtops de l’argó. Aquesta cronologia s’estima entre 4,2 i 3,9 Ma. Atès que els fòssils presenten particularitats exclusives, Meave Leakey i altres col·legues [6] van proposar el 1995, a la revista Nature, la creació de la nova espècie Australopithecus anamensis. El nom específic significa «llac» en la llengua Turkana, i vol recordar que tots els espècimens van ser trobats en sediments que un dia van constituir el fons del llac Lonyumun. Aquest gran dipòsit d’aigua dolça, ja desaparegut, va precedir en el temps a l’actual llac Turkana.
La mandíbula KNM-KP 29281 té un aspecte primitiu, amb la part anterior de la mandíbula fortament inclinada cap enrere i una arcada dental en U, que recorda a les mandíbules d’Australopithecus afarensis i en particular a les trobades en el jaciment de Laetoli. Les canines superiors i inferiors són grans i mostren diferències de mida, a més del desgast característic de la morfologia del complex C/P3 heretada dels simis antropoides. Va poder tenir un dimorfisme sexual potser més proper al dels ximpanzés que al de la nostra espècie. Els mascles van poder pesar fins a un 30% més que les femelles. Es tracta només d’una hipòtesi, molt difícil de contrastar.
En Australopithecus anamensis l’esmalt de les dents és més gruixut que en Ardipithecus ramidus, el que potser revela l’adaptació evolutiva a una dieta més dura i abrasiva. Les espècies fòssils de mamífers (bòvids, antílops, cercopitècids, hienes, rinoceronts, elefants, mustèlids, etc.), peixos i rèptils aquàtics que acompanyen els hominins ens parlen d’ambients oberts de sabanes, amb zones arbrades properes als rius i a l’antic llac. La tíbia humana suggereix bipedisme com es podia esperar d’un hominí d’aquesta antiguitat. El pes de l’individu ha estat estimat entre 47 i 55 quilograms, el que tampoc resulta una sorpresa a jutjar per les dades que es van coneixent de tots els hominins d’aquesta remota època de la humanitat. Finalment, Meave Leakey i els seus col·legues consideren que Australopithecus anamensis és antecessora directa d’Australopithecus afarensis.
Kenyanthropus platyops
Després de la troballa d’Australopithecus anamensis, Meave Leakey i el seu equip van continuar treballant en els voltants del llac Turkana. El 1998 i 1999 es van centrar en la Formació Nachukui, prop de la localitat de Lomekwi, a l’oest d’aquest llac. Aquesta formació geològica conté nombroses capes sedimentàries, diverses de les quals tenen origen volcànic i poden ser datades amb gran precisió mitjançant el mètode de 40Ar39Ar. Meave Leakey i el seu equip de camp van localitzar durant aquests anys fins a 36 restes fòssils humans, la cronologia està molt bé datada entre 3,5 i 3,2 milions d’anys. La major part dels exemplars corresponen a restes mandibulars i dents, com és habitual en tots els jaciments. No obstant això, l’exemplar més representatiu consisteix en un crani (KNM-WT 40000) distorsionat pel pes dels sediments i amb característiques molt peculiars. La combinació de caràcters primitius, compartits amb els australopitecs (incloent un cervell petit), al costat de molts trets presents en espècies del gènere Homo, va portar a Meave Leakey [7] i als seus col·legues a publicar un article a Nature en què es defensava el descobriment d’un nou gènere i espècie: Kenyanthropus platyops.
Es tractava no només d’una volta de rosca més en la diversitat d’hominins del Pliocè, sinó en el reconeixement que a la Gran Vall del Rift van conviure diversos llinatges d’hominins fa entre 4 i 3 Ma. Si el panorama semblava dominat pels australopitecs , l’evolució sembla certament relacionada amb la dels paràntrops, quedava per saber si existien genealogies clarament «dirigides» cap al gènere Homo. El nou gènere descrit per Meave Leakey i els seus col·legues apunta en aquesta direcció, ja que els trets compartits amb Homo (i en concret amb Homo rudolfensis) són majoritaris. Crida particularment l’atenció la cara aplanada del crani KNMWT 40000, que va inspirar el nom de l’espècie i que recorda a la cara plana de KNM-ER 1470, l’holotip d’Homo rudolfensis. L’hominí trobat per Meave Leakey va viure en un ambient humit, cobert de vegetació. Les espècies de bòvids trobades en els sediments de Lomekwi també indiquen la presència de praderies, encara que la major part del paisatge va poder ser de bosc tancat i de boscos de galeria associats als rius que desembocaven llavors al llac Turkana.
Lucy
Molt abans de l’aparició en escena d’Ardipithecus, els investigadors Donald Johanson i Timothy White [8] van dur a terme excavacions i troballes d’enorme rellevància durant la dècada de 1970 a Etiòpia i Tanzània. El 1973 els dos científics van començar a reunir una extraordinària col·lecció de fòssils procedents de la localitat de Hadar (Etiòpia) datats entre 3,3 i 2,9 Ma. Entre aquests fòssils sempre ha destacat l’esquelet d’una femella, que va ser anomenada «Lucy »(protagonista d’una cançó dels Beatles que sonava a l’excavació quan es va trobar el seu esquelet) i considerada durant molts anys com a «mare de tota la humanitat».
Lucy no mesurava més d’un metre d’alçada, el seu pes tot just sobrepassaria els 25 quilograms i el seu cervell no seria més gran que el d’un ximpanzé. En aquells anys, Lucy i una impressionant col·lecció de fòssils d’un mínim de 35 i un màxim de 65 individus representaven els hominins més antics trobats fins llavors. Els seus trets cranials i dentals eren molt primitius i les restes de l’esquelet postcranial (incloent la pelvis de la Lucy) indicaven una incontestable locomoció bípeda.
Alhora, l’equip de Mary Leakey va trobar una altra bona col·lecció de fòssils d’hominins d’una antiguitat encara més gran (3,8-3,6 Ma) a la localitat de Laetoli, a Tanzània. A més, en aquesta mateixa localitat Mary Leakey i Richard Hay van localitzar el 1976 i 1977 les petjades de fins a 20 espècies de mamífers i aus (girafes, babuins, rinoceronts, antílops, elefants, etc.) recognoscibles en una capa ben consolidada de cendres d’origen volcànic procedents de l’erupció del volcà Sadimán. Les cendres es van humitejar amb aigua de pluja i van formar un fangar sobre el qual van quedar les empremtes impreses, més tard cobertes per noves capes de cendra. Entre les petjades de Laetoli destaquen les de dos hominins que caminaven junts. Un d’ells va deixar impressions més profundes, segurament per la càrrega extra que portava a sobre (potser un individu infantil). Per descomptat, els dos caminaven drets i van deixar petjades molt similars a les nostres i diferents, per tant, a les que haguessin deixat els ardipitecs. L’estatura estimada per als dos individus a partir de la empremtes té poca precisió, però no seria inferior a 110 centímetres ni superior a 160 centímetres.
Petjades de Laetoli. Aquestes petjades d’homínid corresponen a tres individus, segurament de l’espècie Australopithecus afarensis, cadascun dels quals seguia les empremtes deixades per un altre individu que caminava just davant d’ell. Laetoli és una de les grans proves que demostren el bipedisme dels australopitecs, ja que no hi ha cap traça ni de mans ni de falanges. A més a més, els peus no presenten el dit gros mòbil com passa amb els grans simis i posseeixen un arc idèntic al dels humans moderns.
Australopithecus afarensis
Johanson i White van realitzar llavors un estudi conjunt de les restes trobades a Hadar i Laetoli. Tot i que les diferències entre les dues mostres eren òbvies, tots dos investigadors van tenir en compte no només la distància temporal i geogràfica, sinó el possible dimorfisme sexual d’aquests hominins. Semblava clar que els mascles eren clarament més alts i pesats que les femelles. Les diferències de forma entre les mandíbules o els maxil·lars podien ser degut simplement a qüestions relacionades amb la mida. En definitiva, el 1978 Johanson i White van proposar al costat de Yves Coppens l’espècie Australopithecus afarensis. Les troballes realitzades a la localitat etíop de Maka, publicades el 1993, semblen omplir el buit morfològic entre els fòssils de Hadar i Laetoli i reforcen la identitat d’Australopithecus afarensis.
Aquesta espècie va viure a l’est d’Àfrica, en un ambient mixt de boscos i sabanes. Les seves adaptacions en l’esquelet postcranial revelen que encara podien enfilar-se amb facilitat, encara que probablement la marxa bípeda fos la manera més habitual de desplaçament.Reconstrucció d’un mascle (esquerra) basat en el fòssil A. L. 444-2, Hadar, Etiòpia, d’uns 3 milions d’anys, i d’una femella (dreta) basat en els fòssils d’A. L. 288-1 i A.L. 417, Hadar, Etiòpia, d’uns 3,2 milions d’anys.

Crani i mandíbula d’AL 444-2.
La troballa d’un esquelet infantil (datat en 3,3 milions d’anys) d’aquesta espècie en la localitat de Dikika, a prop del riu Awash, va resultar molt eloqüent. Aquest fòssil conserva alguns caràcters primitius compartits amb els simis antropoides en l’esquelet postcranial. Malgrat un bipedisme molt avançat, els membres d’Australopithecus afarensis encara van conservar la possibilitat de moure’s amb gran agilitat entre les branques dels arbres quan així ho consideraven oportú. L’entorn de l’Australopithecus afarensis era un ambient de sabana amb sequeres temporals. El seu cervell [9] tenia una mida similar al dels ximpanzés i els seus canins, de bona mida, recorden als del gènere Ardipithecus i al de l’espècie Australopithecus anamensis. És per això que White està convençut de la relació filogenètica entre aquesta última espècie i Australopithecus afarensis que, al seu torn, estaria relacionada amb el llinatge del gènere Paranthropus.
Australopithecus bahrelghazali
És possible que el gènere Australopithecus tingués una distribució geogràfica molt àmplia per bona part d’Àfrica. Michel Brunet i el seu equip de camp han dedicat molt temps a treballar al Txad, lluny de les fructíferes regions arqueològiques i paleontològiques de l’est d’Àfrica. Abans de la troballa de crani de Sahelanthropus, la seva perseverança va tenir un primer premi durant la dècada de 1990, amb la troballa d’un fragment mandibular humà datat en un rang temporal d’entre 3,5 i 3 Ma. Per estimar aquestes dates Brunet i els seus col·legues van utilitzar la informació que ofereix la presència al jaciment de certes espècies de mamífers trobades a Hadar i Laetoli. Brunet va batejar la mandíbula amb el nom d’Australopithecus bahrelghazali, el descobriment va ser publicat el 1995 a la revista Nature [10]. La nova espècie, que seria contemporània d’Australopithecus afarensis, no ha tingut molta influència en els debats sobre evolució humana per tractar-se d’un únic exemplar. No obstant, aquest fòssil mereix ser citat, perquè representa un avantpassat de la humanitat i va ser trobat, com en el cas del gènere Sahelanthropus, a 2.500 quilòmetres de la Gran Vall del Rift, on s’assumeix que va transcórrer tota l’evolució humana. El nom específic al·ludeix a la regió de Bahr el Ghazal, que en llengua àrab significa «riu de les gaseles». Els trets dentals d’aquesta mandíbula suggereixen una relació indubtable amb el gènere Australopithecus i cal esperar que en el futur puguin realitzar noves exploracions en regions tan allunyades de la Gran Vall del Rift.
Fa menys de tres milions d’anys. En els llindars del canvi
Les troballes descrites en els apartats anteriors tenen sempre més de 3 milions d’anys. El període entre 3 i 2 milions d’anys és contradictori.
- d’una banda, tot sembla seguir igual en la nostra evolució.
- de l’altra, s’estava produint un canvi tranquil, però explosiu i determinant del nostre futur.
Anem a repassar troballes de gran transcendència per a la història de l’àmbit de l’evolució humana datades entre 3 i 2 milions d’anys.
Des del punt de vista històric, la troballa de l’anomenat «nen de Taung» va tenir una enorme transcendència en l’àmbit de l’evolució humana. Aquest esdeveniment va suposar un canvi de paradigma en el coneixement dels nostres orígens. El descobriment es va produir per casualitat en 1924, a la pedrera de Taung, a Sud-àfrica, i que feliçment va anar a parar a les mans de Raymond Dart [11]. Els seus coneixements d’anatomia comparada el van fer concloure que el petit crani descobert en aquell lloc no pertanyia a un cercopitècid, com tots els altres fòssils obtinguts en la predera, sinó a un representant del llinatge humà. La morfologia de les dents i la posició basal del foramen magnum indicava clarament que el fòssil de Taung pertanyia a un humà primitiu i que havia caminat dret sobre les dues cames. Dart va nomenar l’espècie Australopithecus africanus, que literalment significa «mico de l’Àfrica austral». Amb aquesta troballa es demostrava per primera vegada que l’origen de la humanitat estava a l’Àfrica i no a Àsia, com havia suggerit Eugene Dubois, però el reconeixement de la seva aportació li va arribar a Dart dotze anys més tard, quan Robert Broom va descobrir l’exemplar de un adult en el jaciment sud-africà de Sterkfontein. Aquesta espècie només s’ha trobat a Sud-àfrica, pel que es pot pensar en un aïllament del llinatge dels australopitecs al sud d’Àfrica fa entre 3 i 2 milions d’anys. Australopithecus africanus va conservar moltes de les característiques primitives dels seus antecessors i amb prou feines va incrementar la mida del seu cervell.
Crani original complet (sense dents superiors i mandíbula) d’un 2,1 milions d’anys d’edat exemplar d’Australopithecus africanus anomenat «Senyora Ples», descobert a Sud-àfrica. Col·lecció del Museu de Transvaal.
La recent datació del jaciment de Sterkfontein, publicada a l’abril de 2015 a la revista Nature per Darryl Granger [12] i els seus col·laboradors, ha deixat més preguntes que respostes. S’ha pogut datar, per primera vegada de manera consistent, un dels nivells de Sterkfontein, que conté l’exemplar STW 573. Aquest fòssil, anomenat «Little Foot» («Peu petit») pel seu descobridor, Ronald Clark, és un dels més complets i conservats de l’espècie Australopithecus africanus. Fins al moment, les datacions per mitjà del paleomagnetisme i el mètode dels isòtops d’urani dels nivells de Sterkfontein havien ofert un ampli marge d’antiguitat entre 4 i 2 milions d’anys. Les investigacions de Darryl Granger i el seu equip per mitjà dels núclids cosmogènics han donat una data de 3,67 milions d’anys. D’aquesta manera, els australopitecs del sud d’Àfrica semblen ser tan antics com l’Australopithecus afarensis. Si és així, aquestes dues espècies d’Australopithecus ja poden competir en igualtat de condicions per la paternitat d’espècies d’hominins posteriors. És més, tradicionalment s’ha considerat que els australopitecs del sud d’Àfrica tenen una forma més generalitzada i estan en millor disposició per ser els ancestres dels primers representants del gènere Homo. Aquest fet potser va ser possible a través dels seus parents de l’est d’Àfrica.
Crani de Little Foot en el lloc de la troballa a la cova de Sterkfontein, novembre del 2006
No obstant, avui per avui, hi molt poques dades per formular una hipòtesi en ferm sobre l’origen del gènere Homo. Es disposa d’una mostra encara molt petita d’exemplars per estar completament segurs que el gènere Homo es va originar fa entre 3 i 2 milions d’anys. Tanmateix, caldrà estar atents a propers troballes. Les investigacions se centraran en aquest període i possiblement arribaran noves sorpreses.
Australopithecus garhi
Una alternativa a les troballes descrits en aquest apartat consistiria a admetre que entre els australopitecs del sud d’Àfrica i els primers Homo va existir alguna forma intermèdia, també relacionada amb el gènere Australopithecus.
Crani reconstruït d’Australopithecus garhi a partir d’elements trobats el 1997 (Awash, regió Afar) de 2,5 milions d’anys. Addis Abeba, Musée national d’Ethiopie.
El crani trobat a nivells sedimentaris de la Formació Bouri (Membre Hata) a la regió mitjana del riu Awash (Etiòpia), datat en 2,5 milions d’anys, podria donar-nos alguna pista pel que fa a aquesta hipòtesi. Berhane Asfaw[13] i altres coneguts paleoantropòlegs han batejat aquest crani amb el nom d’Australopithecus garhi. Per a aquests investigadors, la troballa de l’hominí de Hata va ser un fet inesperat. D’aquí prové el seu nom específic, que en la llengua Afar significa «sorpresa». Asfaw i els seus col·legues pensen que aquesta espècie podria estar relacionada amb Australopithecus afarensis a causa de la seva situació geogràfica, tot i que també reconeixen moltes similituds amb Australopithecus africanus. Les evidències que ens proporciona el crani de Bouri són limitades, però ja ens parlen de la proximitat d’un canvi important en la genealogia humana. Si les eines i els ossos d’animals processats i consumits per hominins trobats en els nivells sedimentaris del Membre Hata van ser obra d’Australopithecus garhi, estaríem a les portes del canvi.
Australopithecus sediba
Una mica més recent en el temps és l’espècie Australopithecus sediba, publicada i descrita amb gran expectació el 2010 a la revista Science per Lee Berger [14] i un conjunt de professionals. L’interès d’aquesta espècie resideix en diversos aspectes. En primer lloc, la publicació descriu dos esquelets molt complets, MHL i MH2, així com la tíbia MH4, trobats a la cova sud-africana de Malapa, a la regió on es localitzen altres jaciments famosos, com Sterkfontein i Swartkrans. L’any 2000, la regió i els seus jaciments van ser declarats Patrimoni de la Humanitat per la Unesco. A més, la cronologia dels dos exemplars s’ha estimat amb notable precisió en 1,98 Ma. És a dir, es tractaria de l’última espècie coneguda d’aquest gènere.
Australopithecus sediba ofereix algunes solucions, però també llança molts interrogants sobre la nostra genealogia. L’aspecte general del crani és més modern que en les altres espècies d’Australopithecus. La nova espècie presenta un veritable mosaic de trets derivats i compartits de manera independent tant amb el gènere Homo com amb el gènere Australopithecus. El seu cervell tenia la mateixa mida que el dels ximpanzés i la seva alçada, proporcions corporals i disseny general de l’esquelet no difereix dels altres australopitecs. L’estudi de les dents donen moltes pistes sobre les relacions filogenètiques d’Australopithecus sediba. És evident que aquesta espècie està emparentada amb Australopithecus africanus, un fet esperable per la proximitat geogràfica, però al mateix temps té indubtables relacions amb Homo habilis, Homo rudolfensis i fins i tot amb Homo ergaster. Aquesta situació resulta paradoxal, perquè en el rang temporal de 3 a 2 milions d’anys tot just es troben indicis del gènere Homo a l’est d’Àfrica, mentre que les proves de la relació entre Australopithecus i Homo es troben a Sud-àfrica. És evident que caldrà construir algun escenari plausible per explicar aquestes relacions filo genètiques en regions allunyades per milers de quilòmetres. No podem oblidar que els primers i clars representants del gènere Homo tenen la mateixa cronologia tant a l’Àfrica com fora del continent (jaciment de Dmanisi) i que l’origen del nostre gènere ha de tenir més de 2 milions d’anys, com suggereix la recent troballa de la mandíbula LD 350-1 de Ledi-Geraru [15]. De l’Australopithecus sediba s’esperen noves dades en els propers anys.
Homo naledi
El jaciment on s’ha trobat Homo naledi (del sesotho naledi, «estrella») va ser descobert pels espeleòlegs aficionats Rick Hunter i Steven Tucker el 2013 a la cova Rising Star a Sud-àfrica, a uns 800 m del jaciment de Swartkrans. El descobriment fou anunciat el setembre del 2015 per l’equip de Lee Rogers Berger[16]. La troballa a la cambra Dinaledi consistia en 15 individus diferents, que comprenen 1.550 exemplars d’ossos. La seva distribució en capes dels ossos suggereix que haurien estat dipositats durant un llarg període de temps, potser fins i tot centúries. Alguns dels ossos s’assemblen als dels humans moderns, mentre que altres ossos són més primitius que el de l’australopitec.
Els fòssils encara no s’han datat però es considera que tenen uns 2,5 milions d’anys d’antiguitat. La posició filogenètica de l’Homo naledi és incerta. Tanmateix les característiques anatòmiques el situen proper als primers Homo: una massa corporal i una estatura similar a la de les poblacions humanes de cos petit (estatura mitjana de 1,50 m i pes de 45 kgr), però amb un volum cranial (entre 465 cm3 i 560 cm3) semblant al dels australopitecs, i amb una morfologia cranial semblant als exemplars més antics del gènere Homo. L’esquelet combina característiques primitives ja presents en els australopitecs amb característiques derivades presents en els primers homínids. La dentició és primitiva i petita. Les mans [17] tenen adaptacions manipuladores humanes. Així mateix són humanes les característiques de cames i peus. No obstant això, el tronc i extrem proximal del fèmur mostren característiques que l’apropen més als australopitecs.
Christoph Zollikofer, de la universitat de Zuric i Tim White, de Berkeley no consideren que es pugui atribuir a una nova espècie, sinó que es podria tractar d’una forma primitiva d’Homo erectus. Per al paleoantropòleg Yves Coppens no és un Homo sinó un Australopitec. Ian Tattersall, del Museu Americà d’Història Natural pensa, també, que els fòssils presenten característiques que els associen més aviat als australopitecs que al gènere Homo. Per a l’antropòleg Jeffrey Schwartz de la universitat de Pittsburgh, els fòssils pertanyen probablement a dues espècies diferents i qüestiona el fet que es tracti d’un ritual funerari.
En resum, els quatre primers milions d’anys de la nostra genealogia ens mostren una gran diversitat d’espècies a Àfrica, que comparteixen un patró comú. Totes elles són bípedes, encara que presenten adaptacions específiques per enfilar-se amb facilitat i suspendre’s de les branques dels arbres. Aquesta possibilitat es va anar perdent a favor d’una locomoció bípeda més habitual a mesura que les selves retrocedien i eren substituïdes per extenses sabanes. La forma general de l’esquelet postcranial sembla mantenir-se a través del temps, amb un tòrax en forma de campana i unes extremitats superiors relativament més llargues que les nostres quan les comparem amb les extremitats inferiors. A més, totes les dades semblen suggerir una alçada baixa en les femelles i una mica més gran en els mascles, com a resultat d’un dimorfisme sexual potser no molt diferent del que mostren avui dia els ximpanzés. El cervell no va augmentar de volum durant tot aquest llarg període de temps i es va mantenir amb una mida similar al dels ximpanzés, tot just superant els 400 centímetres cúbics. És molt possible que la dieta del nostre avantpassat comú amb els ximpanzés tingués un ampli espectre per digerir tot tipus de plantes.
Esquema de les relacions filogenètiques hipotètiques entre les diferents espècies de hominins. La presència de nombrosos interrogants en l’esquema obeeix al fort debat intern dels especialistes, molts dels quals no reconeixen la validesa de certes espècies pobrament representades en el registre fòssil. No obstant això, l’existència real d’aquests fòssils obliga a nomenar-los a l’espera de noves evidències.
Geodiversitat i biodiversitat
Les dues terceres parts de la història del nostre llinatge evolutiu es van desenvolupar a l’Àfrica. Aquest fet no pot estranyar, perquè la majoria de les espècies de mamífers tenen una distribució geogràfica ben definida i molt constant al llarg de centenars de milers o milions d’anys. Les diferents subespècies de ximpanzé comú (Pan troglodytes) tenen el seu hàbitat en diverses regions d’Àfrica, i tot apunta que han romàs en els mateixos llocs durant els últims sis milions d’anys. Els ardipitecs o els australopitecs no van tenir motius per expandir o modificar en gran mesura la seva àrea de distribució i les seves restes fossilitzades probablement es trobaran sempre en les mateixes regions. No obstant això, la geologia ens ensenya que el planeta ha tingut condicions climàtiques molt diferents al llarg de la seva existència. Les espècies continentals són especialment sensibles a aquests canvis i els nostres avantpassats no van ser aliens al que va succeir durant els últims sis milions d’anys. Àfrica, la nostra llar primigènia, va patir les conseqüències d’un canvi climàtic global, que es va accelerar al voltant dels 2,8 milions d’anys. Les investigacions de Peter deMenocal [18] sobre el clima d’Àfrica han estat clau per entendre la història i variabilitat de moltes espècies, incloses les del llinatge humà.
Durant el Pliocè (fa entre 5,3 i 2,6 milions d’anys) el clima de la Terra es va anar refredant i es va tornar més sec i estacional. Aquesta és la situació en la qual encara es troba el nostre planeta. L’Antàrtida es va cobrir amb el seu mantell permanent de gel, tal com avui en dia la coneixem. Àfrica va acusar també aquests canvis i, segons deMenocal, el clima subtropical va estar marcat per oscil·lacions humides i seques. Estàvem encara en l’inici de la formació de les sabanes africanes. No obstant això, els hominins tenien una àrea de distribució molt concreta a les zones més boscoses i amb prou feines van patir les conseqüències d’aquests canvis. El problema va arribar a la fi del Pliocè. Les dates de 2,8, 1,8 i 1 milions d’anys apareixen en totes les investigacions científiques com a moments clau per a la fauna d’Àfrica.
No hi ha acord sobre les raons que van motivar, primer, la successió de canvis climàtics relativament curts i de baixa intensitat i, més tard, l’alternança de fases glacials i interglacials de llarga durada i alta intensitat ocorreguts en l’últim milió d’anys. Els treballs de Milutin Milankovitch sobre la variacions orbitals de la Terra han estat essencials en la teoria sobre l’origen de les glaciacions dels últims tres milions d’anys. La teoria dels cicles de Milankovitch fa referència als canvis orbitals que se succeeixen de tant en tant (les modificacions en l’obliqüitat de l’eix de la Terra, que varien entre 22,1 i 24,5 graus, succeeixen cada 41.000 anys i es corresponen molt bé amb les alternances climàtiques ocorregudes fa entre 2,8 i 1,2 milions d’anys; la raó dels canvis en el clima està en la quantitat de radiació solar que rep el planeta en cada regió: com més gran és l’obliqüitat, més gran és la distinció entre els cicles estacionals).
L’alternança de fases càlides i fredes de 41.000 anys va anar guanyant en intensitat i va aconseguir el seu màxim fa 1,2 milions d’anys. Entre aquesta data i fa 800.000 anys el refredament climàtic de les glaciacions es va accelerar de manera espectacular, començant així les alternances climàtiques de llarga durada i intensitat molt elevada. Altres factors orbitals permeten explicar aquest canvi: l’excentricitat del recorregut de la Terra en la seva òrbita el·líptica al voltant del Sol, el canvis en l’eix de rotació de la Terra, o les oscil·lacions del pla de l’eclíptica que percebem des de la Terra quan observem l’aparent moviment solar al voltant del nostre planeta.
Ara només ens interessa recordar que l’aridesa de l’enorme barrera desèrtica del Sàhara va començar a formar-se coincidint amb l’inici de la nostra genealogia. Aquest fet ens recorda que els efectes climàtics planetaris del Pliocè van començar a influir en el continent africà fa gairebé sis milions d’anys. És molt possible que els primers hominins no estiguessin exposats als canvis que posteriorment arribarien. Per contra la xifra de 2,8 milions d’anys és clau en la nostra història.
El Rift Occidental, també anomenada Albertine Rift, està envoltada per algunes de les muntanyes més altes d’Àfrica, com les muntanyes de Virunga, Mitumba i Range Ruwenzori. Conté els llacs de la Vall del Rift, que inclouen alguns dels llacs més profunds del món (fins 1.470 metres de profunditat al llac Tanganyika).
El continent africà va començar a trencar-se fa uns 30 milions d’anys, durant el Miocè. Des de llavors s’ha produït una fractura de 4.500 quilòmetres de longitud, que comença a l’altura de les costes de l’actual estat de Moçambic i finalitza al Corredor Llevantí, a la vall del riu Jordà, recorrent el fons del mar Roig. Aquesta fractura, amb forma d’Y grega es segueix molt bé per la presència de cadenes de muntanyes, com les Virunga, Mitumba i Ruwenzori, i pels grans llacs, com el Tanganyika, Megadi, Nakuru, Bogoria o Turkana. Aquests llacs representen el preludi de la separació completa d’una gran part de l’est d’Àfrica dins de diversos milions d’anys. No hi ha dubte que el desenvolupament de la Gran Vall del Rift ha estat transcendental en la nostra evolució. La gran majoria dels jaciments d’hominins africans posteriors a 2,5 milions d’anys es localitzen a l’est de la fractura, on el clima ha estat molt diferent al de les regions de l’oest d’Àfrica.
Gran Vall del Rift Africà. Fa 20 milions d’anys, l’escorça terrestre es va començar a afeblir i a separar-se en una fissura que recorre milers de quilòmetres, al llarg de la banya etíop. En aquest punt s’està formant el que dins de milions d’anys serà una dorsal oceànica.
Paràntrops: l’art de sobreviure
Cfr. Tema 17. Paranthropus, del curs en línia Orígens de l’art i evolució humana: l’homo significans. En línia a http://cv.uoc.edu/adf/~04_999_01_u07/llico_17.pdf
Quin va ser l’evolució posterior d’Australopithecus? A partir de 2,6 milions d’anys es produeix un nou tour de force en l’evolució dels homínids africans. En aquest moment els gels s’expandeixen per l’Àrtic, fins llavors lliure d’ells. Per primera vegada des de feia 200 milions d’anys, el planeta entra en una nova era glacial que afecta tant al Pol Nord com al Pol Sud. Fa 2,6 milions d’anys s’inicia l’avanç dels gels permanents sobre Groenlàndia, mentre que al sud les glaceres s’expandeixen sobre la part occidental de l’Antàrtida (la part oriental de l’Antàrtida ja estava gelada des de feia 30 milions d’anys). Després d’aquesta primera glaciació, tot el planeta, i molt especialment el seu hemisferi nord, es veurà afectat per una alternança de polsos glacials que comportaran una successió de fases temperades i fredes. Cada 41.000 anys, coincidint amb un mínim en la inclinació de l’eix terrestre (el que es coneix com obliqüitat de l’òrbita), es produirà una nova extensió de les masses de gel, les quals al seu torn entraran en regressió uns 20.000 anys després (i així successivament). La influència d’aquesta successió de fases glacials i interglacials es va deixar sentir severament en les latituds mitjanes i altes, que van experimentar una important regressió dels boscos temperats, substituïts per una estepa freda semblant a la que avui trobem en àmplies regions de Sibèria. L’important avanç dels gels a l’hemisferi nord va tenir també el seu efecte sobre els ecosistemes africans. Així, a l’Àfrica oriental es va produir una important migració dels boscos cap a les zones equatorials, amb extensió de les sabanes cap al nord i cap al sud. Aquest canvi del clima va donar lloc a hiverns més freds i estius més tòrrids i calorosos. Gramínies i acàcies van passar a dominar el paisatge habitual dels Australopithecus, on abans havien predominat les masses boscoses.
Aquests canvis ambientals tindran una profunda influència en l’evolució dels autralopitecins, de manera que un grup d’ells tendirà a desenvolupar noves adaptacions com a resposta a l’extensió de la sabana. En particular, el que s’observa a partir d’aquest moment és la seva adaptació a una dieta molt més dura i fibrosa, menys omnívora que la dels Australopithecus i basada sobretot en vegetals. Aquesta nova dieta devia estar bàsicament constituïda per llavors, nous i potser arrels, a mesura que els boscos entraven en regressió. Com a resposta a aquest tipus de dieta, sorgeix un nou tipus d’homínids, els paràntrops, els quals, encara que clarament relacionats amb els Australopithecus, mostren clares diferències respecte a ells, sobretot pel que fa al crani. Les dents mastegadores, això és, premolars i molars, es fan cada vegada més grans i es doten d’una gruixuda capa d’esmalt dentari. Al seu torn, els premolars es fan més grans i adquireixen aspecte de molar, per tal de contribuir a la trituració de l’aliment, mentre que els veritables molars es fan progressivament més grans de davant cap enrere (el tercer més gran que el segon, i el segon més gran que el primer).
Aquest imponent aparell dentari ha de ser sostingut per unes grans mandíbules, les quals, al seu torn, necessitaran d’una poderosa musculatura i d’unes àmplies superfícies d’inserció muscular. A conseqüència de la importància adquirida per la musculatura mandibular en aquest grup, la cara s’allarga i els pòmuls es projecten cap endavant, per tal de deixar espai per a uns grans músculs temporals (anomenats així perquè es fixen en els ossos temporals del crani). La conseqüència és una cara llarga i plana, de pòmuls amples i aplanats. Però això no serà suficient, de manera que en els paràntrops, com passa en els actuals goril·les, observem que aquesta potent musculatura temporal envaeix tota la volta craniana, fent fins i tot necessari el desenvolupament d’una cresta, la cresta sagital, que recorre verticalment la part posterior del crani i que permet ampliar la superfície d’inserció d’aquesta musculatura. Com en els goril·les, aquest caràcter era present únicament en els mascles però no en les femelles, ja que, de nou com en els goril·les, els paràntrops mostraven un elevat grau de dimorfisme sexual, amb mascles que podien pesar uns 45 quilos i femelles molt més petites que no arribarien a més de 30 quilos. L’augment de talla que s’observa en els paràntrops té el seu correlat en un lleuger augment de la capacitat craniana, que se situa al voltant dels 500 cc.
Els paràntrops, que sorgeixen fa aproximadament 2,5 milions d’anys amb l’espècie Paranthropus aethiopicus, representen un llinatge molt particular. El 1938 el paleontòleg Robert Broom va batejar amb aquest nom genèric a certs hominins trobats al jaciment sud-africà de Kromdraai. Durant algun temps els espècimens de Paranthropus van ser inclosos per altres especialistes en el gènere Australopithecus. No obstant això, cap a la dècada de 1980 va tornar a imposar-se la idea que aquest llinatge té trets biològics prou diferents com per incloure’ls en un gènere a part.
La resta més antiga de Paranthropus correspon a un crani relativament ben conservat descobert el 1985 per l’equip de Richard Leakey i Alan Walker a la localitat de Lomekwi, a l’oest del llac Turkana. Va ser assignat a l’espècie Paranthropus aethiopicus, una forma fins llavors mal coneguda i que havia estat prèviament definida a la vall del riu Omo, a Etiòpia. Està datat entorn de 2,5 milions d’anys, just en el moment en què les primeres glaciacions s’estenen per l’hemisferi nord. Aquest crani, denominat «crani negre» per la seva pàtina fosca (deguda a alteracions químiques dels sediments que el van preservar fins als nostres dies), presenta ja els trets típics d’un Paranthropus, com és la presència d’una prominent cresta sagital. A més, encara que la seva dentició s’ha perdut, les arrels encara incloses en els seus alvèols parlen d’uns grans molars, similars als observats en altres paràntrops. Ara bé, malgrat les seves característiques derivades en la direcció dels paràntrops, el crani negre presenta un conjunt de trets arcaics que recorden encara a Australopithecus afarensis. Així, la cara és molt prògnata però relativament curta i de perfil còncau, i no llarga i aplanada com en altres espècies de Paranthropus. La seva capacitat cerebral és baixa (uns 410 cc) i el crani allargat, recordant de nou en aquest aspecte a Australopithecus afarensis.
Com en el cas d’Australopithecus, el rang geogràfic de Paranthropus s’estén també fins Sud-àfrica, on el paleontòleg Robert Broom va trobar en 1938 les primeres restes d’aquest gènere, erigint per a ells l’espècie Paranthropus robustus. D’acord amb els espècimens d’aquesta espècie trobats en els jaciments de Swartkrans, Kromdraai i Drimolen, tots ells en l’entorn de Johannesburg, Paranthropus robustus (que va habitar al sud del continent fa entre 2 i 1,5 milions d’anys) presentava grans molars d’esmalt molt gruixut, potents mandíbules i cranis dotats d’una prominent cresta sagital. La seva capacitat craniana se situava entre 500 i 550 cc, estant subjecte a un clar dimorfisme sexual. L’altura estimada dels mascles era d’aproximadament 130 cm, mentre que les femelles havien d’arribar els 110 cm.
Mentrestant, a l’Àfrica oriental, Paranthropus aethiopicus va donar lloc a partir de fa 2,2 milions d’anys a una forma clarament més derivada, Paranthropus boisei (que va viure a l’est d’Àfrica fa entre 2,3 i 1,2 milions d’anys). És en aquesta espècie on la tendència a desenvolupar grans molars arriba al seu màxim, aconseguint proporcions mai vistes en un altre homínid. El crani d’Australopithecus boisei és així mateix el més especialitzat de tots els paràntrops, amb una cara alta i molt verticalitzada, de perfil pràcticament convex en lloc de còncau i pòmuls amples i molt avançats. La seva capacitat craniana és relativament gran per a un paràntrop, entre 475 i 550 cc. Paranthropus boisei, l’últim dels paràntrops, persistirà fins fa uns 1,2 milions d’anys, tal com ho testifiquen les restes de Chesowanja a Kenya i Peninj a Tanzània, extingint-se a continuació, potser a causa de la competència exercida per un altre tipus d’homínids que aleshores poblaven Àfrica oriental.
Randall Susman [19] va arribar a la conclusió que els paràntrops van tenir pinça de precisió per manipular objectes. No obstant això, fins al moment cap arqueòleg els atribueix la capacitat per fabricar eines.
Representació de l’exemplar OH 5 del jaciment d’Olduvai, a Tanzània. Aquest exemplar va ser trobat en 1959 per Mary Leakey i atribuït al gènere Zinjanthropus. Més endavant, va ser inclòs en el gènere Paranthropus i representa l’exemplar tipus de l’espècie Paranthropus boisei. https://www.youtube.com/watch?v=6Y4CMQ82_hQ
El cervell dels paràntrops va créixer en consonància amb un augment moderat de la seva mida corporal. La capacitat de la seva caixa craniana varia d’uns 400 centímetres cúbics en l’espècie més antiga, Paranthropus aethiopicus, i fins a gairebé 550 centímetres cúbics en Paranthropus boisei.
Lectures de referència
Agustí, Jordi; Bufill, Enric i Mosquera, Marina (2012). El precio de la inteligencia. Barcelona. Crítica. (Col. Drakontos). 256 pàgs.
Allman, John Morgan (2003). El cerebro en evolución. Barcelona. Ariel. (Col. Ariel neurociencia). 1ª impr. 240 pàgs.
Arsuaga, Juan Luis i Martínez, Ignacio (2006). La especie elegida : la larga marcha de la evolución humana. Madrid. Ed. Temas de Hoy. 1ª impr. 448 pàgs.
Briones, Carlos; Fernández, Alberto i Bermúdez, José María (2015). Orígenes. El universo, la vida, los humanos. Barcelona. Crítica. Pàgs. 341-469
Campàs, Joan (2014): 15. Els homínids: els preaustralopitecins. Curs en línia: «Els orígens de l’art i l’evolució humana: l’Homo significans». En línia a http://cv.uoc.edu/adf/~04_999_01_u07/llico_15.pdf
Campàs, Joan (2014): 16. Els hominins: els australopitecins. Curs en línia: «Els Orígens de l’art i evolució humana: l’homo significans». En línia a http://cv.uoc.edu/adf/~04_999_01_u07/llico_16.pdf
Campàs, Joan (2014): 17. Paranthropus. Curs en línia: «Els orígens de l’art i evolució humana: l’homo significans». En línia a http://cv.uoc.edu/adf/~04_999_01_u07/llico_17.pdf
Carbonell i Roura, Eudal (2005). Homínidos : las primeras ocupaciones de los continentes. Barcelona. Ariel. (Col. Ariel prehistoria ). 864 pàgs.
Cela-Conde, Camilo i Ayala, Francisco (2011). Senderos de la evolución humana. Madrid. Alianza ed. (Col. Alianza ensayo,188). 7ª impr. 632 pàgs.
Leakey, Richard Erskine (2000). El origen de la humanidad. Barcelona. Debate. (Col. Temas de Debate). 1ª impr. 224 pàgs.
Striedter, Georg F. (2005). Principles of brain evolution. Sunderland MA. Sinauer Associates. 436 pàgs.
Tattersall, Ian (2012). Los señores de la tierra. La búsqueda de nuestros orïgenes humanos. Barcelona. Pasado y Presente. 1ª impr. 352 pàgs.
Notes
[1] Senut, Brigitte; Pickford, M.; Gommery, D.; Mein, P.; Cheboi, K.; i Coppens, Y. (2001). «First hominid from the Miocene (Lukeino Formation, Kenya)». Comptes Rendus de l’Académie des Sciences. París, Sciences de la Terre/Earth and Planetary Sciences Volum 332, Issue 2, 30 de gener del 2001, pàgs. 137-144. En línia a http://www.sciencedirect.com/science/article/pii/S1251805001015294
[2] Veure Orrorin tugenensis a http://www.cnrs.fr/cw/dossiers/Origine/carte/orrorin.htm
[3] Brunet, M. i altres (2002). «A new hominid from the Upper Miocene of Chad, Central Africa». Nature 418, 145-152. En línia a http://www.nature.com/nature/journal/v418/n6894/full/nature00879.html
[4] Haile-Selassie, Yohannes (2001). «Late Miocene hominids from the Middle Awash, Ethiopia». Nature 412, 178-181.
[5] Haile-Selassie, Yohannes (2004). «Late Miocene Teeth from Middle Awash, Ethiopia, and Early Hominid Dental Evolution». Science 303, 1503-1505
[6] Leakey, Meave G i altres (1995). «New four-million-year-old hominid species from Kanapoi and Allia Bay, Kenya». Nature, 376, 565-571.
[7] Meave Leakey i altres (2001). «New hominin genus from Eastern Africa shows middle Pliocene lineages». Nature 410, 433-440.
[8] Johanson, D.C. i White, T.D. (1979). «A systematic assessment of early African hominids». Science 203, 321-330. En línia a http://www.ganino.com/games/Science/science%20magazine%201978-1979/root/data/Science%201978-1979/pdf/1979_v203_n4378/1747757.pdf
[9] William H. Kimbell, Yoel Rak, Donald C. Johanson (2004). The skull of Australopithecus afarensis. Nova York. Oxford University Press. 254 pàgs.
Kimbel, W. H., Johanson, D. C., Rak, J. (1994): «The first skull and other new discoveries of Australopithecus afarensis at Hadar, Ethiopia». Nature 368: 449-451.
[10] Brunet, M. i altres (1996). «The first australopithecine 2.500 kilometres west of the Rift Valley (Chad)». Nature 378, 273-275.
[11] Raymond A. Dart: «Australopithecus africanus: The man-ape of South Africa». Nature.7 de febrer del 1925. 115, 195–199. En línia a http://www.nature.com/nature/ancestor/pdf/115195.pdf
[12] Darryl E. Granger i altres: «New cosmogenic burial ages for Sterkfontein Member 2 Australopithecus and Member 5 Oldowan». Nature, 4 de juny del 2015, 522: 85-88. En línia a http://www.nature.com/nature/journal/v522/n7554/full/nature14268.html
[13] Asfaw, Berhane i altres (1999). «Australopithecus garhi: A new species of early hominid from Ethiopia». Science 284, 629-635. En línia a http://www.indiana.edu/~origins/teach/P314/Bouri.pdf
[14] Berger, Lee R. i altres (2010). «Australopithecus sediba: A new species of Homo-like australopith from South Africa». Science, 328, 195-204.
Berger, Lee R. «Australopithecus sediba and the earliest origins of the genus Homo». Journal of Anthropological Sciences. Vol. 90 (2012), pp. 1-16 En línia a http://www.profleeberger.com/files/Berger_JASs.pdf
[15] LD 350-1 és el nom d’un fragment de mandíbula fòssil, atribuïda a un espècimen d’Homo, que data de fa 2,75-2.800.000 d’anys. Va ser trobada el 2013, al jaciment de Ledi-Geru, regió d’Afar, Etiòpia, per Chalachew Seyoum. Es tracta de la meitat esquerra de la mandíbula inferior d’un adult, amb cinc dents i combina trets trobats en els Australopithecus, amb morfologia derivada observada en el gènere Homo.
[16] Berger, Lee R (i altres): «Homo naledi, a new species of the genus Homo from the Dinaledi Chamber, South Africa», eLife, 2015;4:e09560, 10 setembre 2015. En línia a http://elifesciences.org/content/4/e09560.full
[17] Tracy L. Kivell, Andrew S. Deane, Matthew W. Tocheri, Caley M. Orr, Peter Schmid, John Hawks, Lee R. Berger i Steven E. Churchill: «The hand of Homo naledi». Nature Communications 6: 8431, 6 d’octubre del 2015. En línia a http://www.nature.com/ncomms/2015/151006/ncomms9431/full/ncomms9431.html
[18] deMenocal, Peter B. (1995). «Plio-Pleistocene African climate». Science 270, 53-58. En línia a http://www.indiana.edu/~origins/X-PDF/deMenocal95.pdf
[19] Susman, Randall, L. (1994). «Fossil evidence for early hominid tool use». Science 265, 1570-1573. En línia a http://www.ganino.com/games/Science/science%20magazine%201993-1994/root/data/Science%201993-1994/pdf/1994_v265_n5178/p5178_1570.pdf
Dr. Joan Campàs Aura digital
Estudis d’Arts i Humanitats de la UOC
Curs Som les nostres connexions
Material en format pdf
Tema 0. Presentació i justificació
Tema 1. El cervell ens enganya
Tema 2. El cervell també enganya l’autopercepció del cos
Tema 3. Anatomia cerebral bàsica
Tema 4. Mite: Només utilitzem el 10 % del nostre cervell
Tema 5. Mite: L’homosexualitat (l’hetero, la bi, la trans) és una elecció
Tema 6. Mite: Els fills hereten la intel·ligència dels pares
Tema 7. Mite: Internet i els videojocs són addictius
Tema 8. Mite: El cervell és un ordinador
Tema 9. La memòria
Tema 10. Com definir el «jo»? El cervell i la «joïtat»
Tema 11. L’evolució del cervell (1)
Tema 12. L’evolució del cervell (2)
Tema 13. L’evolució del cervell (3)
Tema 14. L’evolució del cervell (4)
Tema 15. Cervell humà i evolució
Conjunt dels posts amb les introduccions fetes al facebook
Som les nostres connexions