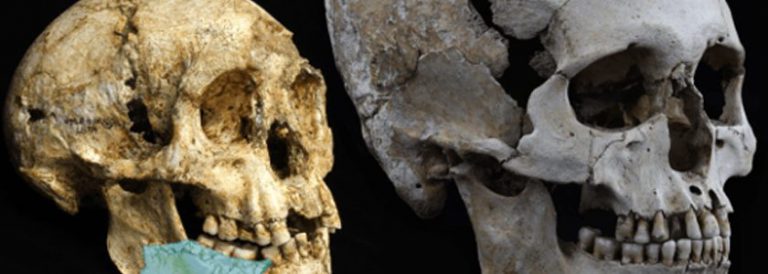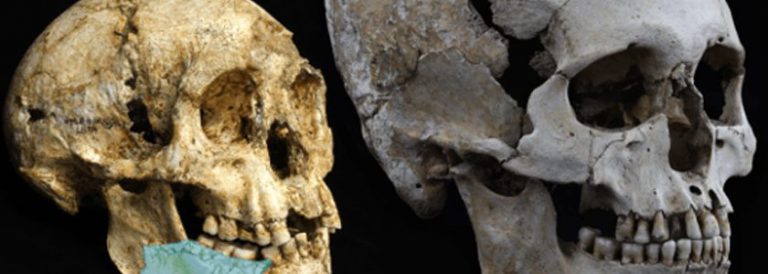
La primera humanitat
Homo habilis
Cfr. Tema 18. Els primers Homo: Homo habilis del curs en línia Orígens de l’art i evolució humana: l’homo significans. Pàg. 2-14. En línia a http://cv.uoc.edu/adf/~04_999_01_u07/llico_18.pdf
Fa una mica més de 2,5 milions d’anys i en paral·lel a l’evolució dels paràntrops, va sorgir a l’Àfrica un nou llinatge humà, que donaria lloc a la nostra pròpia espècie. Molt probablement aquesta nova genealogia es va formar a partir de l’evolució d’alguna espècie del gènere Australopithecus.
Les restes més antigues del nostre gènere es troben en sediments datats entorn a 2,3 i 2,4 milions d’anys, al llarg d’un ampli segment d’Àfrica oriental que inclou Etiòpia, Kenya i Malawi. Aquestes primeres restes, molt fragmentàries, han estat atribuïdes a dues espècies diferents, Homo habilis i Homo rudolfensis.
El gènere Homo es pot reconèixer sense cap mena de dubte en nivells de jaciments amb menys de 2 milions d’anys. En 1964 Louis Leakey, Philip Tobias i John Napier [1] van donar a conèixer a la revista Nature la nova espècie Homo habilis a partir dels espècimens trobats a la gorja d’Olduvai, a Tanzània. Tot i que en les dues últimes dècades diversos investigadors han reclamat que Homo habilis fos considerada com una espècie més del gènere Australopithecus, la veritat és que la força del costum ha impedit que aquesta reclamació tingués massa seguidors. Els arguments esgrimits ja el 1999 per especialistes com Bernard Wood i Mark Collard [2] es basen en el fet ben provat que els membres d’Homo habilis van tenir un creixement i un desenvolupament molt similars als dels australopitecs. Per a aquests investigadors, els petits habilis no havien passat els límits exigibles a una espècie per considerar-la en l’àmbit d’un nou gènere. No obstant això, els espècimens trobats a Olduvai tenien trets distintius que resultaven molt convincents.
La capacitat craniana era d’uns 675 cc, superior a la dels Australopithecus coneguts fins aleshores. Les dents d’Homo habilis eren també diferents dels d’Australopithecus. Així, els premolars eren petits i arrodonits i encara persistia la mateixa polaritat de mida creixent dels molars (el tercer més gran que el segon, el segon més gran que el primer), aquests eren de mida reduïda i esmalt més prim. Malauradament, aquest primer troballa d’Homo habilis no incloïa la cara.
El 1968, de nou a la Capa 1 d’Olduvai va aparèixer un nou crani, OH 24 (o Olduvai Hominid 24), el qual, un cop restaurat, va proporcionar informació sobre la cara i altres detalls del crani. La capacitat craniana era d’uns 600 cc, inferior a l’exemplar de 1964, però de nou superior a la de qualsevol Australopithecus. El més sorprenent d’OH 24, però, va ser que, a part de la seva capacitat craniana, la forma de la cara, baixa i còncava, recordava en molts aspectes a la d’Australopithecus africanus. Les concomitàncies entre Homo habilis i Australopithecus van més enllà de la seva anatomia facial, afectant també a l’esquelet. Així, el 1987 Don Johanson i Tim White van donar a conèixer la troballa a la Capa 1 d’Olduvai d’un esquelet parcialment complet que ells van atribuir a Homo habilis. L’esquelet, conegut com OH 62, incloïa un paladar, fragments del crani, restes de l’húmer i del radi, part del fèmur i de la tíbia així com altres peces menors. L’anàlisi d’OH 62 va demostrar que la proporció entre braços i cames era molt semblant a la d’Australopithecus afarensis, amb membres anteriors relativament més llargs que en altres espècies posteriors. Això sembla indicar que Homo habilis encara retenia les capacitats arborícoles dels seus avantpassats australopitecins. Mentre que les restes de la Capa 1 d’Olduvai se situen al voltant de 1,9 milions d’anys, el rastre d’Homo habilis s’ha pogut seguir a Kenya i Etiòpia fins a prop de 2,4 milions d’anys enrere.
Gràcies a les dades del registre arqueològic, sabem que alguna cosa important estava succeint a l’Àfrica en el lapse de temps que transcorre entre fa 3 i 2 milions d’anys.
Mode 1. Tecnologia olduvaiana
Cfr. Tema 3. Evolució tecnològica en el Paleolític. Referències etnoarqueològiques de l’hàbitat del curs en línia Orígens de l’art i evolució humana: l’homo significans. Pàgs. 1-6.En línia a http://cv.uoc.edu/adf/~04_999_01_u07/llico_3.pdf
Les eines de pedra trobades a la base de la seqüència geològica de la gorja d’Olduvai tenen una antiguitat de 1,8 Ma. Es tracta de percussors, ascles simples o retocades i els nuclis de les que es van extreure. La tecnologia és relativament senzilla, però implica un canvi substancial pel que fa a les eines que utilitzen els ximpanzés de manera ocasional. La tecnologia Olduvaiana, com se la coneix des de la dècada de 1960, implica la transformació de la matèria primera amb una finalitat i mitjançant una acurada i elaborada planificació. Els ximpanzés utilitzen pals i pedres. En aquesta tasca hi ha per suposat un pla determinat. Però els humans vam arribar més lluny en planificar l’elaboració de les eines, amb l’objectiu d’obtenir una major eficàcia. Aquesta elaboració implica una transformació de les propietats de la matèria i suposa un salt cognitiu en aparença digne de tenir en consideració.
Les investigacions en jaciments de Kenya (Turkana) i Etiòpia (Omo) van envellir fins a 2,3 milions d’anys l’aparició de la tecnologia lítica, encara que les dades més antigues van ser aportats per les excavacions de Sileshie Semaw [3] i procedeixen de les localitats OGS-6 i OGS -7 de la regió de Gona, un afluent del riu Awash, a Etiòpia. Les datacions en aquestes localitats arriben fins a 2,6 milions d’anys. La majoria de les eines estan realitzades amb materials d’origen volcànic i inclouen els mateixos tipus que a Olduvai. La selecció dels materials denota una planificació complexa, mentre que la seqüència de cops compassats amb un percussor per aconseguir la peça desitjada suggereix gran habilitat. És evident que la troballa d’instruments de pedra encara més antics és només qüestió de temps. A més, crida l’atenció el fet que la tecnologia Olduvaiana no hagués experimentat canvis substancials durant gairebé un milió d’anys a l’Àfrica. Els humans que van exportar aquesta tecnologia fora d’Àfrica, que avui dia es coneix com Mode 1, tampoc van dur a terme innovacions en la producció d’eines. La talla de la pedra va romandre a Euràsia amb un grau de complexitat gairebé similar fins fa uns 800.000 anys.
Els dubtes sobre l’espècie que va fabricar les eines de Gona i d’altres jaciments de Kenya i Etiòpia persistiran mentre no es reforci la hipòtesi que la mandíbula L.D. 350-1 de Ledi-Geraru pertany al gènere Homo.
Alguns arqueòlegs suggereixen que els membres d’alguna espècie d’Australopithecus i fins i tot de Paranthropus van ser els responsables de la fabricació dels instruments de pedra més antics de la genealogia humana. El treball publicat per Matthew Skinner [4] apunta en aquesta direcció. Les investigacions de Skinner parteixen de diversos fets ben coneguts. L’os pateix canvis importants durant el seu creixement i al llarg de la vida. Aquests canvis tenen una base genètica, però també un fort component ambiental. El treball al qual se sotmeten els ossos determina en part el seu aspecte extern i intern. Els ossos responen a l’exercici físic i a la funció que es realitzi amb assiduïtat. S’ha observat que la densitat de la trabècula òssia [5] dels metacarps dels australopitecs i paràntrops és similar a la dels ximpanzés. Aquestes dades confirmen que els nostres avantpassats eren capaços de grimpar, agafant-se a les branques amb una força enorme. Però la distribució de la trabècula òssia dels metacarps en australopitecs i paràntrops és molt similar a la nostra i diferent a la dels ximpanzés. Ja que l’orientació de les trabècules està condicionada per la direcció de la força que s’exerceix sobre elles, hi ha la possibilitat d’inferir la funció que realitzaven els ossos de les mans dels nostres avantpassats. De l’estudi de Skinner i els seus col·legues es desprèn que tant els australopitecs com els paràntrops van ser capaços de fabricar instruments. La conclusió de Skinner podria explicar la presència d’eines en èpoques tan remotes, en absència d’espècies confirmades del gènere Homo. Potser els australopitecs van ser els primers a iniciar l’era de la tecnologia.
Sònia Harmand [6] ha relatat la troballa de diversos centenars d’eines al jaciment de Lomekwi 3, pròxim al llac Turkana, a Kenya. Aquest jaciment s’ha datat en 3,3 milions d’anys, de manera que l’aparició de la tecnologia tindria arrels encara molt més profundes. Potser no ens quedi més remei que admetre d’una vegada per totes que la tecnologia no ha estat exclusiva del gènere Homo, sinó d’Australopithecus i potser d’algun altre gènere de la nostra genealogia. L’equació: cervell gran = tecnologia, manejada fins al moment, no té per què ser correcta. El «salt mental» que suposa colpejar una pedra per obtenir un tall tallant pot semblar gegantí, o potser no ho és tant. La necessitat de viure en espais oberts i de consumir amb més freqüència la carn d’animals va poder ser un al·licient perquè les espècies d’Australopithecus donessin aquest salt cognitiu. Sembla, doncs, que ens trobem davant d’un canvi de paradigma i que el concepte d’ésser humà haurà de patir una profunda revisió.
Ens preguntem ara per l’ús d’aquests instruments. Les eines apareixen associades a restes fòssils de grans herbívors que mostren les marques dels talls dels ganivets de pedra. Exemples d’interès apareixen al jaciment de Bouri, a la regió del curs mitjà del riu Awash, amb una cronologia de 2,5 milions d’anys.
Des de fa diverses dècades, científics com Lewis Binford, Jean de Heinzelin [7], Pat Shipman, Yolanda Fernández-Jalvo o Manuel Domínguez-Rodrigo s’han ocupat d’estudiar les marques que s’observen en la superfície dels ossos fòssils i, en particular, a prop de les articulacions. Amb la introducció de les tècniques de microscòpia electrònica es va poder caracteritzar amb enorme detall la morfologia d’aquestes marques. Sens dubte, es tracta dels estigmes que van deixar els hominins en actuar sobre els cadàvers dels animals per tallar trossos de carn, seccionar tendons o colpejar les canyes dels ossos llargs i extreure’n la medul·la. En general, els animals processats corresponen a diferents espècies de mamífers de gran talla: elefants, hipopòtams, rinoceronts, cavalls, cérvols, gaseles, bisons, etc. Per exemple, en un treball recent de 2013, Mohamed Sahnouni [8] i el seu equip de col·laboradors han descrit evidències molt clares del processament de cadàvers de grans mamífers en el jaciment d’El-Kherba, prop d’Ain Hanech, al nord d’Algèria. Aquest jaciment té una antiguitat de 1,8 Ma i prova que la pràctica del consum de carn de mamífers de gran talla s’havia estès per totes les regions africanes on vivien els humans.
De seguida ens sorgeixen diversos dubtes. ¿S’alimentaven els australopitecs amb la carn d’aquests animals ?, o van ser representants desconeguts del gènere Homo els autors d’aquests festins carnívors? La possibilitat que els australopitecs consumissin proteïnes d’origen animal és una hipòtesi molt plausible, perquè els ximpanzés també ingereixen una certa quantitat de carn, incloent la d’altres espècies de primats. La pràctica esporàdica de canibalisme també s’ha observat en els ximpanzés. Una altra qüestió és l’accés a aquests grans mamífers. És probable que l’empitjorament climàtic obligués els australopitecs a aconseguir recursos carnis en mitjans oberts. O potser van ser els primers i desconeguts membres del gènere Homo els que van incrementar la ingesta de carn. A més, queda el dubte plantejat fa diverses dècades sobre la possibilitat que els hominins no fossin grans caçadors, sinó carronyers actius o passius. La mida dels australopitecs o dels habilins (Homo habilis) no sembla el més apropiat per caçar elefants o rinoceronts, ni fins i tot actuant en grups ben organitzats. És per això que alguns científics van plantejar la possibilitat que els hominins aconseguissin la carn d’aquests animals ben aprofitant les carcasses abandonades, bé disputant els recursos a altres carronyers.
La hipòtesi que Lewis Binford [9] i Richard Potts [10] van plantejar en els anys vuitanta del segle XX ha estat objecte de fortes debats. Ells i molts altres experts han postulat que els nostres primers avantpassats del gènere Homo només van tenir la possibilitat d’accedir als cadàvers de grans mamífers matats per altres animals. En altres paraules, els habilins van ser catalogats com carronyers de les sabanes. Aquesta visió dels nostres avantpassats ha romàs com una hipòtesi robusta durant anys. El paper dels carronyaires no té «bona premsa», tot i que voltors, aufranys, trencalòs, hienes i multitud de petits organismes s’encarreguen de netejar l’entorn de matèria orgànica en putrefacció. Els voltors i les hienes representen l’exemple millor conegut per tots com carronyers actius, que es nodreixen de cadàvers complets o devorats a mitges per predadors. En el primer cas, els carronyers poden matar per si mateixos aprofitant la debilitat per malaltia o vellesa d’algun animal. En altres casos, els observadors del cel estaran alerta d’alguna presa parcialment devorat, a la qual acudiran tant ells com altres carronyers. En totes les ocasions, uns i altres participen d’una rivalitat activa per aconseguir algun mos de l’animal caigut.
Ja que molts investigadors són incapaços d’imaginar a un petit grup de habilins disputant el menjar a voltors i hienes, s’han proposat variacions de la hipòtesi del «hominí carronyer». Alguns han optat per assignar un paper de carronyers passius als nostres avantpassats. Aquesta hipòtesi alternativa assumeix que els nostres ancestres estaven atents a les matances dels tigres dents de sabre (Homotherium), els enormes canins dels quals els impedien devorar per complet a les seves preses. La resta seria consumit pels petits habilins i altres espècies de la nostra genealogia. La clau resideix en què els tigres dents de sabre segurament caçaven en boscos relativament tancats, on les seves preses no quedaven exposades a l’agudesa visual dels carronyers voladors.
Sobre la possibilitat que haguéssim pugnat de manera activa per petites quantitats de carn adherides als ossos de les preses es poden plantejar alguns dubtes. No només per la dificultat que comportaria la lluita aferrissada per aconseguir un mos, sinó perquè molta de la carn trobada en les sabanes estaria en estat de descomposició. Els voltors tenen un aparell digestiu perfectament adaptat per consumir carronya, malgrat el seu alt contingut en bacteris de gran toxicitat, com el mateix àntrax (Bacillus anthracis) o representants dels gèneres Fusobacterium i Clostridium. Altres vertebrats i nosaltres mateixos podríem morir amb la ingesta de matèria orgànica infectada per aquests microorganismes. Sorgeix llavors una pregunta inevitable: ¿vam tenir els primers representants del gènere Homoaquest tipus d’adaptacions protectores en l’aparell digestiu? I si és així, per què les vam perdre?
No podem descartar que els nostres avantpassats tinguessin la fortuna d’accedir a preses mortes de manera natural o de restes d’animals caçats per altres predadors, però encara amb carn i greix en condicions de certa salubritat. Segurament, però, també vam ser capaços de capturar altres vertebrats, inclosos els espècimens juvenils, així com d’enriquir la dieta amb multitud d’invertebrats en les ribes de llacs i rius, que s’afegirien amb menor perillositat al nostre menú. La hipòtesi de l’hominí carronyer resulta llavors massa simple, perquè implica diversos passos evolutius complexos i contradictoris.
De moment tots els experts estan d’acord que la capacitat tecnològica està ben desenvolupada en els espècimens d’Homo habilis d’Olduvai, com ens indica l’estudi dels ossos de la mà. Els canvis anatòmics a la mà són imprescindibles per manipular objectes amb precisió. Aquestes modificacions impliquen el desenvolupament dels músculs adductors i flexors del polze, el primer dit de la mà, així com del múscul flexor llarg del polze, que s’origina en el radi i s’insereix a la base de la falange distal d’aquest dit. En els ximpanzés no existeix aquest múscul, i la seva funció és assumida per un tendó (flexor digitorum profundus). Aquests canvis anatòmics són molt evidents i afecten el desenvolupament i força muscular del polze i l’increment de la seva grandària per aconseguir oposar-se als altres dits de la mà i, en particular, al dit índex. Sense aquestes modificacions anatòmiques només és possible realitzar la pinça de pressió, que permet subjectar objectes amb la mà, però no la seva manipulació precisa. Per aquest motiu la possibilitat de la fabricació d’estris de pedra per part dels australopitecs requereixi la demostració, més enllà de tot dubte raonable, que aquests hominins tenien pinça de precisió o, si més no, una capacitat més desenvolupada que la dels ximpanzés per manipular objectes.
En definitiva, la fabricació sistemàtica d’eines de pedra, el consum de carn de grans mamífers i la pinça de precisió semblen haver-se originat fa una mica més d’2,5 milions d’anys. Els habilins d’Olduvai tenien un cervell ni més ni menys que un 40-50% més gran que el dels australopitecs. És evident que aquest salt no es va produir de la nit al dia, sinó que es va poder gestar al llarg del període que hi ha entre 2,5 i 1,8 milions d’anys.
Si busquem una explicació plausible a l’increment de la mida cerebral, haurem de pensar tant en els nutrients que necessitem perquè el cervell pugui créixer com a la selecció positiva d’un òrgan «molt car» de mantenir en termes energètics. Per descomptat, la llet materna té tots els ingredients necessaris per al desenvolupament del cervell dels seus fills. Però les mares han d’accedir abans als nutrients adequats, com els àcids grassos omega 3 o l’àcid docosahexaenoic (DHA).
D’altra banda, els hominins van haver de modificar el seu comportament per aconseguir aliments menys predictibles que els que es troben a l’interior dels boscos. Els animals es mouen i són perillosos. Si aquests animals van constituir una part substancial de la dieta dels hominins és evident que els nostres ancestres van haver de millorar les seves prestacions cognitives.
Homo rudolfensis
Cfr. Tema 18. Els primers Homo: Homo habilis del curs en línia Orígens de l’art i evolució humana: l’homo significans. Pàgs. 14-15. En línia a http://cv.uoc.edu/adf/~04_999_01_u07/llico_18.pdf
La segona espècie arcaica del gènere Homo, Homo rudolfensis, presenta caràcters bastant diferents d’Homo habilis. Pitjor coneguda que l’anterior, la peça que ens ha proporcionat major informació sobre ella és un crani bastant complet, que respon a la sigla KNMER 1470, descobert el 1972 per l’equip de Richard Leakey a la vora de llac Turkana, a Kenya. El més notable d’aquest crani, que conserva bona part de la cara i tot el neurocrani (la part del crani que allotja el cervell), és la seva capacitat craniana, de més de 750 cc, notablement més gran que la de les restes atribuïdes a Homo habilis (al voltant de 650 cc). La cara és plana i molt allargada, diferent de la dels representants posteriors del gènere Homo. Pel que fa a la dentició, encara que al crani ER 1470 li falten les dents, la forma i dimensions dels alvèols dentaris indiquen una dentició robusta més pròxima a la dels Australopithecus (i, fins i tot, els parántropos) que a la del propi Homo habilis. L’edat de KNM-ER 1470 se situa al voltant dels 1,9 milions d’anys però, d’acord amb un fragment de mandíbula procedent de Malawi, el rang temporal d’Homo rudolfensis pot estendre’s fins fa gairebé 2,4 milions d’anys.[11]
Així doncs, fa entre 2,4 i 2,3 milions d’anys es produeix l’eclosió d’un nou tipus d’homínid, representat per una o dues espècies (alguns autors consideren que Homo habilis i Homo rudolfensis, corresponen en realitat a una mateixa espècie ). Ara bé, on es troben les arrels del gènere Homo? ¿A partir de quina mena de australopitecí es va desenvolupar? Encara que ara per ara no hi ha una resposta definitiva a aquestes preguntes, al llarg de les últimes dècades s’han proposat diversos candidats potencials. En el cas d’Homo rudolfensis, la publicació en 2001 d’un nou crani procedent de la Formació Lomekwi (Kenya) per part de Meave G. Leakey i altres col·laboradors va oferir una versió aventurada sobre el seu origen. El crani, encara que format per múltiples fragments i molt deformat a la part posterior, presentava una associació de caràcters inèdita per a un fòssil l’edat del qual es va xifrar en 3,5 milions d’anys. Molts d’aquests caràcters eren molt arcaics i congruents amb un fòssil contemporani d’Australopithecus afarensis, com era la seva baixa capacitat craniana i el seu petit canal auditiu extern. Per contra, el que va sorprendre als autors de la troballa va ser l’aspecte de la seva cara, plana i projectada anteriorment per sota de l’orifici nasal, amb pòmuls alts i aplanats. Donada aquesta particular associació de caràcters, els descobridors del crani de Lomekwi van decidir erigir un nou gènere i espècie, Kenyanthropus platyops, per la nova troballa. Ara bé, segons els signants de l’article, la morfologia facial de Kenyanthropus recordava la d’altres formes més recents com el crani ER-1470, assignat a Homo rudolfensis. Per tant, tot i ser prop d’un milió i mig d’anys més recent, les arrels d’Homo (o Kenyanthropus) rudolfensis caldria rastrejar 3,5 milions d’anys enrere en ple apogeu d’Australopithecus afarensis. Ara bé, malgrat la seva cara aplanada i prògnata, hi ha altres caràcters que permeten diferenciar Kenyanthropus platyops d’Homo rudolfensis com és la seva dentició de mida reduïda (més fins i tot que la d’Australopithecus afarensis), que casa malament amb la suposada dentició d’ER -1470, molt més robusta.
D’altra banda, el 1999, Tim White i el seu equip van descriure una nova espècie d’Australopithecus, Australopithecus gahri que, per a ells ocuparia una posició intermèdia entre Australopithecus afarensis i les primeres espècies del gènere Homo [12]. La troballa realitzada a la localitat de Hata Bouri a Hadar (Etiòpia), incloïa nombroses restes poscranials però també un fragment cranial que inclou bona part del neurocrani i del paladar. La cara d’Australopithecus ghari presenta una morfologia més arcaica que la d’Australopithecus africanus i les formes primitives del gènere, més propera en aquest sentit a la del seu probable antecessor afarensis. D’aquest últim, però es diferencia per alguns caràcters més avançats que afecten la dentició, i que indicarien una certa tendència a desenvolupar molars grans i gruixuts. La seva capacitat cranial és baixa, havent estat estimada en uns 450 cc. Per a Tim White i el seu equip, Australopithecusghari se situa en el lloc adequat i en el moment precís per a ser considerat com l’avantpassat directe dels primers representants de nostre gènere.
A pocs quilòmetres de Hata Bouri, a Gona, s’ha trobat la més antiga evidència d’instruments lítics, datats en uns 2,5 milions d’anys. Qui va ser l’autor de les eines de Gona? Segons Tim White, Australopithecus gahri casaria com el perfecte candidat per ser l’autor de les mateixes, donada la coincidència cronològica i geogràfica entre Hata Bouri i Gona. Aquesta assumpció es veu reforçada pel descobriment de marques de descarnament sobre alguns dels ossos de la fauna acompanyant Australopithecus gahri. Les marques de descarnament apareixen com una sèrie d’incisions paral·leles que s’observen en aquells ossos que han estat descarnats mitjançant algun tipus d’útil tallant. Així doncs, si es confirmés l’autoria d’aquesta espècie pel que fa a la indústria lítica de Gona i a les incisions observades en els ossos de Hata Bouri, tindríem que la capacitat per a elaborar instruments lítics hauria precedit a l’expansió cerebral que es detecta en els primers representants del gènere Homo.
El 2010, un nou candidat a avantpassat directe d’Homo va saltar a la palestra. Es tracta d’una nova espècie, l’Australopithecus sediba, trobat per l’equip de Lee Berger [13], de la Universitat de Witwatersrand, a Johannesburg, en una cavitat càrstica a uns 50 quilòmetres al nord d’aquesta ciutat. La troballa està compost per dos esquelets parcials que pertanyen a una dona jove d’uns vint anys i a un adolescent d’uns dotze anys, aquest últim representat per un crani en excel·lent estat de conservació que revela a un Australopithecus pròxim a Australopithecus africanus però amb caràcters més derivats. La forma de la cara recorda a la d’Homo habilis, però la capacitat del crani, encara que una mica més elevada que la dels seus predecessors, és encara petita, al voltant dels 420 cc. Els braços i, sobretot la mà, indiquen una bona aptitud per enfilar-se pels arbres. Els dits són més curts que en altres espècies i el polze és més llarg, el que, segons els seus descobridors, hauria habilitat a Australopithecus sediba per fabricar eines. La pelvis de l’exemplar femení és ja semblant a la que es troba en les formes evolucionades d’Homo, mentre que els ossos del peu mostren una barreja de caràcters arcaics i derivats. Australopithecus sediba, per tant, estaria ben adaptat tant per a passejar per la sabana com per sortir-se’n entre els arbres. En aquest sentit, aquesta espècie apareixeria com la perfecta baula intermèdia entre Australopithecus africanus i els primers representants del gènere Homo. El problema és que les restes d’Australopithecus sediba estan datats en uns 1,9 milions d’anys i són, per tant prop de mig milió d’anys més recents que els primers Homo d’Àfrica oriental. Fins i tot, aquesta espècie d’Australopithecus seria coetània dels primers representants del nostre gènere que van sortir d’Àfrica. Ara bé, el que sí és possible és que l’hipotètic avantpassat d’aquests primers Homo fos una forma molt pròxima a Australopithecus sediba, encara que cronològicament anterior.
Preparats per al primer viatge
Un esdeveniment transcendental d’aquesta època està relacionat amb la primera migració ben documentada dels hominins fora del continent africà. Aquesta expansió va succeir a través del Corredor Llevantí seguint el curs de la Gran Vall del Rift.
Dmanisi. Homo georgicus
Curiosament, les evidències més completes sobre l’anatomia i manera de vida d’aquesta primera humanitat no procedeixen d’Àfrica sinó que se situen fora d’aquest continent, al sud del Caucas. Així, la localitat georgiana de Dmanisi ha proporcionat el conjunt més complet d’homínids corresponent a una forma que amb prou feines sobrepassa a Homo habilis en la major part dels seus caràcters [14]. Es tracta de la primera dispersió humana més enllà d’Àfrica. Fins al moment en aquest extraordinari jaciment, a més d’una variada fauna de grans i petits mamífers, s’han extret les restes de diversos homínids, corresponents tots ells a una mateixa població, incloent cinc cranis més o menys complets, quatre mandíbules, un fèmur, una tíbia, un peroné, diversos húmers, vèrtebres, costelles i diversos ossos de la mà i el peu pertanyents a un individu juvenil i a altres tres individus adults [15].
La primera troballa es va produir el 1991, amb motiu d’una excavació paleontològica dirigida per Leo Gabounia i Abesalom Vekua als voltants d’un castell medieval proper al llogaret de Dmanisi. Aquest lloc està situat a uns 90 quilòmetres al sud de Tbilisi, la capital de la República de Geòrgia. Després de gairebé nou anys d’incertesa sobre l’antiguitat del jaciment i la posició filogenètica de les restes humanes, la comunitat científica va reconèixer finalment que els cinc esquelets trobats fins al moment en el jaciment de Dmanisi tenen una cronologia de 1,8 milions d’anys.
Els homínids de Dmanisi, assignats a l’espècie Homo georgicus, presentaven una baixa capacitat craniana, oscil·lant entre els 775 cc de la calota D2280 i els 600 cc del crani D2700. Es tracta de valors que s’enquadren dins dels límits de variabilitat de les poblacions més arcaiques d’Homo, com Homo habilis i Homo rudolfensis. D’acord amb el crani D2700, un dels més ben conservats del conjunt, aquests primers pobladors d’Euràsia mostraven una cara baixa i còncava, caracteritzada a nivell del maxil·lar per un acusat prognatisme, el que confereix a aquesta part del crani un perfil còncau i no convex com en altres homínids posteriors. Es tracta d’individus petits, de talla reduïda, tal com ho demostra el crani D2700, tot just 15 cm d’un extrem a un altre del crani. Tot i que la dentició era, en general, reduïda, mostraven unes canines relativament grans i prominents. Una nova mandíbula, apareguda l’any 2000, va permetre fer-se una idea més clara de la variabilitat poblacional a la població de Dmanisi. Aquesta segona mandíbula estava més completa que la primera de l’any 1991 i mostrava una sèrie de característiques que la feien molt diferent de l’anterior. Així, aquesta mandíbula excedia la mida dels més grans exemplars atribuïts al gènere Homo, com la mandíbula 3 del jaciment nord-africà de Ternifine (o Tighenif), d’uns 700.000 anys d’antiguitat, convertint-se així en la resta més gran d’aquest tipus atribuït al nostre gènere. Les branques mandibulars eren molt altes, mostrant que el seu posseïdor havia d’haver presentat una cara així mateix molt alta: l’alçada de les dues branques ascendents era de gairebé 10 cm. D’altra banda, malgrat el seu acusat desgast, va ser possible comprovar que les incisives havien estat de mida molt petita, especialment les centrals. Al seu torn, les canines haurien estat comparativament grans, a jutjar per les seves potents arrels. Quant als premolars, tot i que només es conservaven en bon estat els primers, va ser possible observar que aquests mostraven dues arrels en lloc d’una, un caràcter certament molt arcaic. Els molars eren grans i massius, d’esmalt gruixut (prop de 2 mm de gruix en alguns punts). Però el més sorprenent va ser la polaritat de mida d’aquests molars: el tercer molar, molt gran, sobrepassava en talla al segon, el qual al seu torn estava més desenvolupat que el primer.
Aquesta barreja de caràcters, entre arcaics i altres més derivats cap a les espècies posteriors del gènere Homo, s’observa així mateix a nivell de l’esquelet. De l’anàlisi de les diverses parts esquelètiques trobades (diversos húmers, fèmur, tíbia, peroné, diversos ossos de la mà i del peu) es dedueix que els homínids de Dmanisi van ser individus de petita talla, d’entre 40 i 50 quilos i una alçada que se situaria entre els 140 i 150 cm. De cintura cap amunt, diferien de les espècies posteriors del gènere Homo en una sèrie de caràcters arcaics, com l’absència de torsió en el cap de l’húmer, sent comparables en aquest sentit als australopitecins. En la major part de representants del gènere Homo, nosaltres inclosos, el cap de l’húmer presenta una típica torsió que fa que els palmells de les nostres mans mirin més aviat cap al cos. Això no succeeix ni en ximpanzés ni en goril·les, ni tampoc en els australopitecins i les formes arcaiques d’Homo, on les mans tendeixen a mirar cap endavant. És possible que aquesta funció es relacioni amb una millor aptitud per viure als arbres, i que aquesta interpretació sigui extensible també als homínids de Dmanisi. Ara bé, si observem la seva anatomia de cintura cap avall, el panorama és molt diferent. Així, el fèmur és relativament llarg, de manera que la proporció entre braços i cames és ja típicament «moderna», comparable a la nostra i la major part d’espècies del gènere Homo. Així mateix, el peu mostra un arc ben desenvolupat, una característica que ja es troba en els representants d’Homo habilis.
Ara bé, la major sorpresa proporcionada pel jaciment de Dmanisi i que llança una nova llum sobre el comportament d’aquests arcaics integrants de la primera humanitat es va produir durant les campanyes de 2003 i 2004. En aquests anys, i de manera consecutiva, va sortir a la llum un nou crani amb la seva corresponent mandíbula (sigles D3444 i D3900, respectivament). Aquest crani no diferia substancialment dels anteriors, encara que es tractava d’un individu plenament adult. La seva capacitat cerebral era igualment petita, al voltant dels 650 cc i per regla general, presentava característiques similars als anteriors. Ara bé, a diferència d’ells, el nou crani i la seva mandíbula no tenien dents (excepte una resta d’incisiva a la mandíbula). L’individu, probablement senil o malalt, havia perdut les seves dents feia alguns anys, de manera que els alvèols dentals havien desaparegut, obliterats per l’os, un caràcter molt patent a la mandíbula, d’una extraordinària primesa [16]. Un fenomen semblant s’havia notat en el crani de la Chapelle-aux-Saints, a França. Però aquest crani corresponia ja a un neandertal datat en poc més de 50.000 anys, la capacitat cerebral del qual era ja similar a la nostra. Per contra, l’individu desdentat de Dmanisi corresponia a una població de fa 1,7 milions d’anys i el cervell tot just excedia en una mica més de 200 cc al d’un ximpanzé. En absència de fruits i davant la necessitat de mantenir una dieta amb una fort component carni, això indica que l’individu senil de Dmanisi va ser alimentat activament pel grup i mantingut amb vida durant dos o més anys. En el context de les dures condicions de vida dels homínids de principis del Plistocè, quin sentit biològic tenia mantenir a un individu que no podia valer-se per si mateix i que ja havia complert amb les seves funcions reproductores? No ho sabem, però, en qualsevol cas, Dmanisi demostra que el comportament cooperatiu estava ja arrelat en els orígens mateixos del gènere humà fa prop de dos milions d’anys.
Fins a mitjans de l’última dècada del segle passat es postulava que aquesta primera expansió fora d’Àfrica havia estat protagonitzada fa un milió d’anys per humans d’elevada alçada, capaços de recórrer llargues distàncies en poc temps, i amb un cervell de dimensions entorn a els 900 centímetres cúbics. Aquesta hipòtesi va haver de ser rebutjada gràcies als descobriments de Dmanisi.
Els primers humans a sortir fora d’Àfrica, doncs, no complien les exigències que es van pensar en un principi. Ni eren massa alts ni el seu cervell era massa gran. Però, llavors, ¿per què es va produir aquesta expansió dels humans fora d’Àfrica?
Hipòtesis
1. És possible, com pensen alguns investigadors, que la diversitat d’hominins en els ecosistemes de la Gran Vall del Rift produís una competència per l’espai i els recursos, el que va obligar a certes poblacions a moure’s cap al nord. Recordem que molts especialistes reconeixen fins a tres espècies del gènere Homo entorn a 1,9 – 1,8 milions d’anys: Homo habilis, Homo rudolfensis i Homo ergaster. L’espècie Australopithecus sediba, trobada, descrita i definida a Sud-àfrica, no hauria competit amb aquestes espècies tot i que la seva cronologia s’ha xifrat en gairebé 2 milions d’anys: el seu hàbitat se situa a diversos milers de quilòmetres de l’est d’Àfrica.
Homo rudolfensis segueix sent una incògnita, perquè l’únic exemplar ben conegut és el crani KNM-ER 1470, trobat en 1972 per l’equip de Richard Leakey a la riba oriental del llac Turkana. Aquest crani té una capacitat craniana de 750 centímetres cúbics i una cara gran i aplanada, diferent de la que presenten els exemplars d’Homo habilis. Potser KNM-ER 1470 va pertànyer a una espècie diferent, com reconeixen molts investigadors, i el seu nínxol ecològic va ser diferent al dels habilins. Finalment, ens queda l’espècie Homo ergaster, que alguns paleoantropòlegs prefereixen incloure en Homo erectus. Deixarem de banda aquest debat taxonòmic.
2. Potser aquesta diversitat no va ser el factor decisiu per explicar la primera expansió dels hominins fora d’Àfrica. És probable, en canvi, que la dieta fos el motiu que induís una major mobilitat dels grups humans. Si el seu menjar es movia, ells també haurien de fer-ho. Algunes espècies d’herbívors van emigrar cap al nord de la Gran Vall del Rift i els humans possiblement van seguir els seus passos. No obstant això, quan es realitza un estudi de la fauna trobada a Dmanisi amb prou feines es troben espècies d’origen africà.
3. Sense descartar aquesta última possibilitat, encara ens queda una tercera hipòtesi. És possible que en aquest temps es produís un creixement demogràfic dels hominins, així com una escassetat de recursos coincidint amb algun període de major aridesa. En aquest possible escenari va sorgir la necessitat d’ampliar l’hàbitat dels hominins, que van haver de moure’s per territoris de major extensió. No cal pensar en una ràpida migració per explicar la sortida d’Àfrica. A raó de tan sols un quilòmetre anual es pot arribar a la serralada del Caucas en 6.000 anys des de les regions ocupades en l’actualitat per l’estat d’Etiòpia. Totes les dades paleoecològiques apunten que la regió de Dmanisi representava llavors un d’aquests reductes de clima benigne i recursos abundants, que van caracteritzar l’hemisferi nord durant el Miocè.
La investigadora Marie Antoinette de Lumley [17] ha batejat als hominins de Dmanisi amb el nom específic d’Homo georgicus basant-se en l’exemplar de major grandària (mandíbula D 2600), les dimensions són significativament més grans que les de les mandíbules més petites. Els cranis corresponents a aquestes mandíbules tenen notables similituds amb els de l’espècie Homo ergaster i reminiscències indubtables d’Homo habilis. En canvi, el crani 5 (format pels exemplars D 2600 I D 4500) té una mida notablement superior i un desgast dental molt peculiar i diferent al dels altres individus. Aquest crani resulta una veritable incògnita pel seu descomunal aparell mastegador. Potser la diversitat que apareix a Dmanisi està reflectint la pròpia diversitat d’Àfrica. El temps s’encarregarà de donar una resposta.
La segona humanitat
Homo ergaster
Cfr. Tema 19. El poblament d’Euràsia: l’Homo ergaster/erectus del curs en línia Orígens de l’art i evolució humana: l’homo significans. Pàgs. 1-8. En línia a http://cv.uoc.edu/adf/~04_999_01_u07/llico_19.pdf
Homo ergaster és la denominació que molts utilitzen per anomenar les restes fòssils africanes compreses entre 1,9 i 1,4 milions d’anys. Aquesta espècie inclou exemplars tan coneguts com KNM-ER 992, KNM-ER 3733, KNM-ER 3883 i KNM-WT 15000.
Hi ha la possibilitat que l’espècie arribés a viure a l’Àfrica fins fa un milió d’anys. La seva capacitat craniana s’estima entre 800 i 900 centímetres cúbics i l’estatura d’alguns individus va poder superar amb comoditat més d’un metre i mig. Es pot dir que amb Homo ergaster es van aconseguir les proporcions corporals de la nostra espècie.
Pel que fa a l’Àfrica, al voltant de fa uns 1,6 milions d’anys també trobem les restes d’un nou tipus d’homínid, Homo ergaster, que recorda en molts aspectes a l’asiàtic Homo erectus. El nostre coneixement d’Homo ergaster és molt més complet que el d’Homo erectus, gràcies sobretot al descobriment el 1984 d’un esquelet adolescent molt complet, popularment conegut com «el noi del Turkana», o «Turkana boy» (descobert el 23 d’agost del 1984 per l’investigador Kamoya Kimeu, membre de l’equip de paleoantropòlegs dirigit per Richard Leakey). Procedent dels sediments de Nariokotome, situats al marge oriental del llac Turkana, a Kenya, el «noi del Turkana» tenia en morir una edat d’uns dotze anys i mesurava 160 cm d’alçada, tot i que d’haver arribat a estat adult probablement hauria arribat als 180 cm. El sorprenent del cas és que l’esquelet del noi del Turkana mostra, a partir del cap, unes proporcions típicament modernes, amb cames llargues i braços relativament curts, molt semblants en molts aspectes a les dels actuals pobladors del llac. Els vestigis anatòmics que encara vinculaven a Homo habilis a una vida arborícola, amb els seus braços relativament llargs, han desaparegut ja a Homo ergaster, que, d’aquesta manera, esdevé una forma d’homínid típicament adaptada a la sabana oberta (com els actuals massai) . A diferència dels paràntrops o del mateix Homo habilis, Homo ergasterva ser capaç d’abandonar la vida sedentària dels seus predecessors i recórrer llargues distàncies a través de la sabana oberta, a la recerca d’aigua i recursos, tal com fan avui els moderns massai i altres tribus de la zona del Turkana. És possible també que Homo ergaster representi la transició des d’unes poblacions purament carronyeres cap a altres que haurien experimentat ja la caça sistemàtica. I, de fet, això és el que suggereixen les restes lítiques associats a aquesta espècie. Així, fa 1,7 milions d’anys es produeix l’eclosió d’un nou tipus de cultura, l’Acheuliana (o Mode 2), que comportarà una revolució en la manera de fabricar i utilitzar els estris lítics. La base d’aquesta revolució cultural la constitueix un instrument bàsic, el bifaç, que caracteritzarà culturalment a la humanitat en el següent milió i mig d’anys.
Cfr. Tema 3. Evolució tecnològica en el Paleolític. Referències etnoarqueològiques de l’hàbitat del curs en línia Orígens de l’art i evolució humana: l’homo significans. Pàgs. 6-10.En línia a http://cv.uoc.edu/adf/~04_999_01_u07/llico_3.pdf
Podem inferir que potser va ser en aquest moment quan, per primera vegada, la humanitat va adquirir la típica nuesa que avui caracteritza la nostra espècie. Davant la necessitat de recórrer llargues distàncies sota el sol, és molt possible que fos llavors quan aquests homínids africans van desenvolupar l’eficaç sistema de sudoració que avui permet refrigerar el nostre cos davant l’excés de calor. La pèrdua del pèl hauria permès estendre per gairebé tot el cos el sistema de glàndules sudorípares i sebàcies que és característic de la nostra espècie i que ens diferencia dels nostres germans antropomorfs.
Quin va ser l’origen d’Homo ergaster? Podem teoritzar que, atès que fa uns 1,8 milions es detecta a l’Àfrica una nova volta de rosca cap a una progressiva aridesa, amb expansió de la sabana herbàcia i regressió de les masses boscoses, algunes poblacions d’Homo habilis s’haurien adaptat a les noves condicions ambientals, augmentant el volum del seu cervell i modificat l’aparell locomotor. Ara bé, aquestes mateixes modificacions s’observen gairebé simultàniament en els Homo erectus asiàtics. Podem pensar que a partir de la humanitat de Dmanisi es va produir una evolució en paral·lel en ambdós continents. Ara bé, hi ha un escenari alternatiu més senzill, segons el qual els canvis en la capacitat cerebral i en l’aparell locomotor s’haurien produït poc després de Dmanisi en alguna regió d’Àsia central. Des d’allà, aquesta segona humanitat s’hauria expandit en dues direccions: cap a l’est, fins a arribar als confins d’Àsia, i de nou cap a Àfrica, derivant en el que coneixem com a Homo ergaster, que per a alguns no deixa de ser sinó un Homo erectusafricà. Així doncs, després del primer «out of Africa» que representa la població de Dmanisi, s’hauria produït un moviment al revés, un autèntic «into Àfrica», el qual explicaria la sobtada presència d’Homo ergaster en aquell continent. Hi ha evidències que fan versemblant aquest últim escenari. Per exemple, fa al voltant d’1,6 milions d’anys es produeix la coexistència a l’Àfrica oriental d’autèntics Homo ergaster al costat de les últimes poblacions d’Homo habilis, el seu teòric avantpassat africà. Tot i que la coexistència d’una espècie i el seu ancestre dins d’una mateixa àrea no és impossible, l’esquema evolutiu es fa molt més senzill si admetem que Homo ergaster procedeix d’Homo georgicus i no directament d’Homo habilis.
Sens dubte, el més interessant d’aquests hominins resideix en el gran pas que va suposar l’inici de la prolongació del temps del creixement i el canvi en el model de desenvolupament. La clau d’aquesta troballa es va publicar el 1985 a la revista Nature, quan Timothy Bromage i Christopher Dean [18] van ser capaços d’esbrinar la durada del temps del desenvolupament d’espècies extingides mitjançant l’estimació del temps de formació de les seves dents. El temps de formació de la corona i l’arrel de les dents forma part del creixement somàtic de tot l’organisme, incloent per suposat el creixement cerebral. Encara que s’ha postulat que el cervell és l’òrgan que regula tot el nostre creixement, una mena de «marcapassos» de tot el que succeeix, no hi ha dades concloents que recolzin aquesta hipòtesi. En tot cas, podem dir que tot el nostre organisme creix i es desenvolupa de manera organitzada, jerarquitzada i ordenada, amb més o menys lentitud, i que les velocitats de desenvolupament de cada part no tenen per què ser les mateixes. A partir de cert moment, els hominins van alentir tot el seu desenvolupament. Les dents fòssils, que es conserven a milers en els jaciments, van ser la clau per entendre com i quan va succeir aquest canvi tan important en la nostra evolució.
Bromage i Dean van establir les bases per a entendre l’últim milió i mig d’anys de la nostra evolució. Fins el 1985 tots els experts acceptaven que els australopitecs tenien una infància prolongada i una llarga fase d’adolescència. Però la prolongació de la infància, que alguns experts divideixen en infància (0-2 anys) i infantesa (2-6 anys), va començar fa relativament poc temps, durant l’evolució d’espècies com Homo ergaster. L’adolescència també va poder iniciar-se en aquesta espècie, però no es va acabar de forjar de manera definitiva segurament fins a dates molt més recents. Els australopitecs, els paràntrops i els habilins creixien i es desenvolupaven en el mateix temps i cadència que els ximpanzés. D’aquí les seves similituds biològiques.
Per entendre què ha passat amb el creixement cerebral durant la nostra evolució ens podem fixar en les dades obtingudes per Tomoko Sakai i els seus col·laboradors [19] sobre el creixement del cervell en ximpanzés i humans. El nostre cervell creix a gran velocitat durant la gestació, que en certs moments arriba als 33 centímetres cúbics setmanals, i només disminueix quan s’acosta el moment del part. Gràcies a aquest patró de creixement sortim del ventre matern amb una mida cerebral propera als 400 centímetres cúbics; és a dir, naixem amb la mida del cervell a la que arriben els ximpanzés adults. En aquests primats, la velocitat de gestació màxima és de 15 centímetres cúbics setmanals, que baixa fins als 4,1 centímetres cúbics en les setmanes anteriors al part. En conseqüència, el cervell dels nounats està al voltant de 150 centímetres cúbics. Per entendre què va passar amb el cervell d’Homo habilis tan sols hem de pensar en algun canvi genètic dels molts que sens dubte s’estaven generant i sotmetent a la selecció natural. Aquest canvi va permetre incrementar la velocitat de creixement del seu cervell durant la gestació. El canal del part d’Homo habilis era prou folgat com per permetre donar a llum a cries amb un cervell una mica més gran, potser al voltant dels 230 centímetres cúbics.
Aquestes dades per als nadons d’Homo habilis les trobem a les investigacions de Jeremy de la Silva i Julie Lesnik [20]. Aquests científics van estudiar la mida del cervell de nadons i adults en diverses espècies de primats catarins (els primats d’Àfrica i Euràsia), inclosos els éssers humans. De manera sorprenent, totes les dades es van ajustar perfectament a una recta de regressió: la mida de cervell del nounat i de l’adult estan relacionades i aquesta relació es pot expressar mitjançant una fórmula matemàtica. El cervell dels nounats d’Homo sapiens, tot i ser molt voluminós, és l’esperat per un primat d’aquest grup. Tampoc en aquest aspecte som especials ni diferents a altres espècies, com els macacos, els babuins o els ximpanzés. Ja que tots els nostres ancestres també eren primats catarins, només cal tenir la mida del cervell de l’adult de qualsevol espècie per a realitzar una raonable estimació del cervell dels seus nounats. No obstant això, la corba de regressió obtinguda per De Silva i Lesnik no és una recta del tipus Y = a + bX, sinó que quan augmenta la mida del cervell del nounat, encara ho fa més la mida del cervell de l’adult. Per exemple, el cervell dels ximpanzés nounats (uns 150 centímetres cúbics) representa aproximadament el 37% de la mida del cervell de l’adult, mentre que en l’Homo sapiens el cervell dels nounats (uns 380 centímetres cúbics) representa aproximadament el 28% de la mida del cervell dels adults. Aquest fet està sens dubte relacionat amb la necessitat de no augmentar massa la mida de cervell dels nadons i evitar així problemes durant el part. El consegüent augment de la mida cerebral es pot aconseguir després d’aquest procés, durant la vida extrauterina.
Per exemple, es coneix des de fa molts anys que la taxa de creixement cerebral en Homo sapiens es manté elevada fins que arribem al primer any de vida. Assolim el 90% de la mida cerebral de l’adult cap als cinc anys d’edat i gairebé el 100% quan complim sis anys. En els ximpanzés, el 90% de la mida cerebral s’aconsegueix cap als quatre anys i el 100% cap als cinc anys. En definitiva, tot i que disposem d’un any més per al creixement cerebral que els nostres cosins germans, aconseguim un cervell tres vegades més gran que el seu gràcies a una velocitat de creixement molt superior tant durant la gestació com en els cinc primers anys de vida extrauterina.
Tot i que sabem que no existeix un paral·lelisme perfecte entre desenvolupament cerebral i desenvolupament dental i/o somàtic, no hi ha dubte que els neandertals (Homo neanderthalensis) i altres espècies anteriors, els cervells dels quals van sobrepassar els 1.000 centímetres cúbics, es van aproximar a un model de creixement i desenvolupament molt similar al nostre. Les fases d’infantesa i adolescència, tan importants per al nostre desenvolupament cognitiu, segurament es van apropar a la seva plenitud fa mig milió d’anys. En aquests trets biològics, els humans de fa mig milió d’anys potser no eren tan diferents de nosaltres.
Homo erectus
Cfr. Tema 19. El poblament d’Euràsia: l’Homo ergaster/erectus del curs en línia Orígens de l’art i evolució humana: l’homo significans. Pàgs. 9-17. En línia a http://cv.uoc.edu/adf/~04_999_01_u07/llico_19.pdf
Des del sud del Caucas, aquesta primera humanitat va iniciar un llarg trajecte que l’ha dut en molt poc temps, als confins de l’Àsia tropical a milers de quilòmetres de distància. Efectivament, molt poc després o gairebé al mateix temps que Dmanisi, trobem possibles evidències físiques i culturals d’aquesta presència humana en llocs tan remots com Java [21] o l’interior de la Xina [22]. És el cas dels jaciments de Modjokerto en Java o Longgupo, a Xina datats en gairebé 1,8 milions d’anys. Durant aquest recorregut els arcaics homínids de Dmanisi van patir una profunda transformació. El seu cervell es va engrandir fins a sobrepassar els 800 cc. La volta craniana es va allargar alhora que sobre les òrbites oculars es van desenvolupar unes gruixudes estructures òssies, anomenades arcs supraciliars, que van haver d’actuar com una mena de visera sobre els ulls. Aquest nou tipus d’homínid, diferent de l’anterior, és conegut com Homo erectus i el seu registre es perpetua a Àsia fins fa menys de 100.000 anys.
La primera expansió dels hominins fora d’Àfrica va ser seguida, doncs, d’una migració molt ràpida cap a l’est. Els humans degueren arribar en poc temps a les illes de l’actual estat d’Indonèsia, potser passant per terra ferma durant una fase glacial. Els descensos del nivell del mar en aquestes regions durant els períodes glacials unien tot l’arxipèlag formant l’anomenada regió de Sunda. Els descensos del nivell del mar per acumulació del gel al continent van deixar pas franc als moviments dels hominins fins a l’illa de Java. Amb l’increment del nivell del mar durant els períodes interglacials, els hominins quedaven aïllats durant mil·lennis. L’aïllament poblacional pot permetre explicar la gran diversitat de formes d’hominins a l’arxipèlag d’Indonèsia. Aquesta diversitat va portar de cap als experts durant anys, que van arribar a proposar diversos gèneres i espècies per al registre de hominins del Plistocè d’aquesta regió. Després de molta controvèrsia, la comunitat científica va acceptar que la diversitat de formes d’hominins trobats a Sunda podria ser agrupada sota la denominació d’Homo erectus, amb una història que comença fa almenys 1,6 milions d’anys i acaba fa uns 50.000 anys, amb els humans trobats al jaciment de Ngandong. Els experts estan d’acord a acceptar que la diversitat dels humans trobats en aquest llarg període es deu a l’evolució in situ d’una mateixa espècie, potser subdividida en diverses subespècies, i als perllongats períodes d’aïllament.
Un bona part dels experts en evolució humana han utilitzat el nom Homo erectus per denominar a la major part dels fòssils trobats a l’Àfrica i Euràsia durant el Plistocè. Curiosament, va ser un ornitòleg alemany, Ernst Mayr, qui el 1950 va proposar agrupar la majoria dels fòssils humans del gènere Homo a l’espècie Homo erectus. Tot i que en anys posteriors s’han descrit noves espècies del gènere Homo, l’esperit de la proposta de Mayr va ser tremendament influent. En l’actualitat, i a propòsit de l’estudi dels cranis del jaciment de Dmanisi, s’ha arribat a proposar que tots els fòssils africans del gènere Homo del Plistocè Inferior, incloses les espècies Homo habilis i Homo rudolfensis, haurien de ser incloses en Homo erectus. D’aquesta manera, Homo erectus cobriria una diversitat insòlita per a qualsevol espècie, a més d’una gran distribució geogràfica i una durada temporal de gairebé dos milions d’anys. Aquesta diversitat inclouria morfologies cranials molt diferents, algunes amb cares primitives i altres amb cares molt similars a la d’Homo sapiens (Zhoukoudian) o cervells d’una mida entre 600 i 1.200 centímetres cúbics. Per descomptat, una altra bona part dels experts no està d’acord a incloure a tants i tants hominins en Homo erectus.
L’homo floresiensis
Cfr. Tema 20. Homo antecessor, rhodesiensis, heidelbergensis i floresiensis del curs en línia Orígens de l’art i evolució humana: l’homo significans. Pàgs. 10-18. En línia a http://cv.uoc.edu/adf/~04_999_01_u07/llico_20.pdf
L’anàlisi genètica de dues restes fòssils trobades al jaciment de la cova de Denisova, localitzada a les muntanyes Altai de Sibèria, ha aportat dades molt interessants sobre les relacions entre les poblacions del passat. La capa sedimentària en què van aparèixer les dues restes pot tenir entre 50.000 i 30.000 anys. Aquests fòssils mostren relacions genètiques amb els neandertals, amb els humans de la Sima de los Huesos de la serra d’Atapuerca i amb Homo sapiens. Les dues restes, una falange i un molar d’aspecte molt arcaic, van ser «sacrificades» per obtenir el DNA que contenien. Ja que només s’han trobat aquests dos fòssils humans a la cova de Denisova, ens preguntem si va valer la pena el sacrifici. És més, de no haver-se dut a terme aquesta anàlisi, les dues restes haurien passat inadvertides i haguessin acabat al calaix d’Homo erectus. No obstant això, els experts especulen amb la necessitat de proposar una nova espècie, de la qual ara no hi ha ni un sol fòssil. Són les paradoxes de l’evolució humana.
En contrast amb aquesta història, els genetistes no han pogut recuperar DNA de les restes trobades el 2003 al jaciment de la cova de Liang Bua de l’illa de Flores. Aquesta illa va quedar sempre separada del continent de Sunda, tot i durant els descensos més importants del nivell del mar. Flores va quedar aïllada d’Euràsia durant milers d’anys i es va convertir en un veritable «laboratori evolutiu» per a les espècies que van habitar l’illa, com els elefants nans del gènere Stegodon o els llangardaixos gegants, similars als dracs de l’illa de Komodo. L’esquelet LB1, de 18.000 anys d’antiguitat, va ser clau perquè els seus descobridors donessin nom el 2004 a l’espècie Homo floresiensis. L’alçada d’aquell humà, que havia arribat a l’estat d’adult, era d’uns 100 centímetres i el seu cervell tot just superava els 400 centímetres cúbics. Per aquest motiu, molts arqueòlegs van dubtar de la capacitat cognitiva d’Homo floresiensis per fabricar les eines trobades al jaciment i l’ús intencionat de foc.
Les excavacions d’anys posteriors han arribat a obtenir nombroses restes humanes addicionals. La datació dels dipòsits sedimentaris de Liang Bua es xifra entre 90.000 i 13.000 anys d’antiguitat. Tot i ser tan recents, el DNA d’aquests humans no ha pogut ser seqüenciat i analitzat. Les condicions tropicals de l’illa de Flores degraden amb enorme rapidesa el material genètic, que hagués ajudat a desxifrar l’enigma creat per aquesta troballa. Els defensors que Homo floresiensis representa la conseqüència d’una nanisme evolutiu assumeixen que aquests humans procedeixen d’una població d’Homo erectus, potser com la trobada en el jaciment de Dmanisi. Si és així, aquests experts haurien d’admetre un aïllament de 1,6 milions d’anys a l’illa de Flores i que l’espècie trobada hauria de ser inclosa com a subespècie a Homo erectus floresiensis, per no entrar en contradicció amb els defensors de l’enorme extensió geogràfica i temporal d’aquesta espècie. No obstant això, els nans de Flores són molt particulars en els seus trets cranials i es consideren una de les diverses excepcions que van sorgint a mesura que es produeixen descobriments.
També existeixen treballs que inclouen els nans de Flores a la nostra espècie. Aquestes poblacions haurien patit algun tipus d’anomalia genètica o malaltia, que els va portar a presentar un cervell i una alçada extremadament petits. Peter Obendorf [23], per exemple, suggereix el cretinisme o hipotiroïdisme congènit, amb retard mental associat, a causa d’una manca de iode i seleni en la dieta. D’altres, com Robert Eckhardt [24], sostenen que l’esquelet LB 1 va pertànyer a un individu amb síndrome de Down, ja que els individus amb aquesta anomalia genètica tenen un mateix volum encefàlic en aquesta regió del planeta. Altres científics, com Anita Rauch [25] i el seu equip, postulen que els nans de Flores patien nanisme microcefàlic i osteodisplàsic de tipus II, que no produeix cap retard mental. En fi, tot apunta que la polèmica seguirà en el futur, mentre no es pugui trobar una mica de DNA sense degradació ni contaminació.
LB1 en tres vistes diferents per il·lustrar la asimetria facial. Les diferències en les arquitectures facials laterals esquerra i dreta són evidents, i il·lustren anomalies en el creixement de LB1.
Com a resum d’aquest apartat podem dir que no tots els humans del Plistocè han estat inclosos en Homo erectus. Fins i tot els experts més fonamentalistes i menys oberts al debat admeten altres espècies en aquest període i en altres regions, com veurem en el següent capítol.
La segona sortida d’Àfrica
Fa entre 800.000 i 600.000 anys el nostre planeta va experimentar una nova transició climàtica, de conseqüències molt més dures que l’anterior de fa 2,6 milions d’anys. En aquest moment s’inicia una sèrie de polsos glacials de característiques diferents als anteriors i en els quals encara estem immersos. Això polsos glacials es caracteritzaran per temperatures encara més baixes que les aconseguides en anteriors fases fredes i per l’extensió de grans masses de gel (o inlandsis) sobre el nord d’Europa i Amèrica del Nord. L’anterior ciclicitat climàtica de 41.000 anys és substituïda per una altra de 100.000 anys, controlada aquest cop per la forma de l’òrbita (més o menys excèntrica) i no per la obliqüitat de l’eix de la Terra. A diferència dels cicles de 41.000 anys, durant els quals s’alternaven fases glacials i interglacials de durada semblant (d’uns 20.000 anys cadascuna), en els nous cicles glacials s’alternen fases fredes molt llargues (d’uns 80.000 anys) i fases interglacials relativament breus, d’uns 20.000 anys (nosaltres ens trobem actualment a la meitat d’un d’aquests interglacials).
Com a conseqüència, una estepa freda s’estendrà per bona part d’Europa i Àsia. Grans mamífers adaptats a aquestes noves condicions passaran a dominar els ecosistemes terrestres, com ara els mamuts d’estepa (Mammuthus trogontherii), els rinoceronts d’estepa (Stephanorhinus hemitoechus) els cavalls d’aspecte modern (Equus mosbachensis) els bisons (Bison priscus) o els primers bous salvatges (Bos primigenius). A diferència de les altes latituds, a l’Àfrica aquests polsos glacials no es traduiran en descensos significatius de la temperatura però sí en fases d’acusada aridesa, amb noves expansions de la sabana seca i, molt probablement, també del desert. Pel camí es van quedar els últims paràntrops, desapareguts abans de fa un milió d’anys. Tot i que el registre d’homínids africans en el període comprès entre fa un milió i 600.000 anys és molt pobre, hi ha indicis que Homo ergaster va prosseguir la seva evolució, d’acord amb diverses restes cranials relativament ben conservats procedents de la Capa 2 d’Olduvai, de Buia, a Eritrea, i de Bouri, a l’Awash mitjà a Etiòpia [26]. Totes aquestes restes retenen caràcters que els fan reminiscents d’Homo erectus, però al mateix temps mostren ja caràcters més avançats cap a un nou tipus d’homínid, com és la possessió d’una caixa craniana que s’eleva verticalment.
Homo rhodesiensis
Cfr. Tema 20. Homo antecessor, rhodesiensis, heidelbergensis i floresiensis del curs en línia Orígens de l’art i evolució humana: l’homo significans. En línia a http://cv.uoc.edu/adf/~04_999_01_u07/llico_20.pdf, pàgs. 5-6
I efectivament, a partir de fa uns 600.000 anys trobem les primeres evidències d’una nova forma humana, caracteritzada per una elevada capacitat craniana, que pot arribar fins als 1.400 cc. Aquesta espècie va ser ja reconeguda a l’Àfrica a principis del segle XX, quan a la localitat de Broken Hill (avui Kvabue, a Zàmbia, l’antiga Rhodèsia britànica) es va extreure un crani ben conservat al costat d’altres restes esquelètiques corresponent a un homínid de port modern, amb un cervell de prop de 1.300 cc i un crani de parets verticals. Les òrbites oculars eren grans i estaven coronades per gruixuts arcs supraciliars discontinus, que no formaven la prominent visera dels anteriors Homo erectus. La cara era gran i ampla i estava projectada cap endavant. En mans del paleontòleg britànic Arthur Smith Woodward, aquest investigador va proposar una nova espècie per allotjar a l’exemplar de Broken Hill, Homo rhodesiensis. Amb posterioritat, noves restes han incrementat la mostra d’aquesta espècie a l’Àfrica, com és el cas de crani de Bodo, a Etiòpia [27].
Homo antecessor versus homo heidelbergensis
Cfr. Tema 20. Homo antecessor, rhodesiensis, heidelbergensis i floresiensis del curs en línia Orígens de l’art i evolució humana: l’homo significans. En línia a http://cv.uoc.edu/adf/~04_999_01_u07/llico_20.pdf, pàgs. 1-4
Però el més rellevant d’aquest nou tipus d’homínid és que a partir de fa uns 600.000 anys, les restes d’una espècie molt similar, Homo heidelbergensis, es troben per bona part d’Europa. Així, fa uns 500.000 trobem evidències anatòmiques a Alemanya (una mandíbula en Mauer, prop de Heidelberg) i al sud d’Anglaterra (un fragment de tíbia a Boxgrove), ambdues situades a uns 50° de latitud nord. Gairebé de la mateixa edat, o potser fins i tot anterior, és l’extraordinària col·lecció d’aquesta espècie del jaciment de la Sima de los Huesos, a Atapuerca. Restes d’aquesta espècie es troben també a Talteüll, a la Catalunya francesa (fragments cranials i esquelètics datats en 450.000 anys). Una mica més recents, entorn a 400.000 i 300.000 anys, són els fragments cranials de Bilzingsleben, a Alemanya, i de Swanscombe, a Anglaterra. Encara més recent, confirmant la continuïtat del poblament humà a Europa al llarg de tot el Plistocè mitjà, és el crani molt complet de Steinheim, d’uns 250.000 anys d’antiguitat, i el crani de Petralona, a Grècia, datat en uns 200.000 anys . Així doncs, després de la primera sortida d’Àfrica protagonitzada per la gent de Dmanisi, tot indica que per segona vegada en la història humana un segon homínid va arribar a aventurar-se fora del nostre continent matriu. Ara bé, a diferència dels anteriors, Homo heidelbergensis va aconseguir el que cap espècie anterior d’homínid havia aconseguit: endinsar-se en les fredes i inhòspites estepes i boscos d’Europa central.
Des de fa menys d’una dècada, l’equip investigador d’Atapuerca ve defensant la hipòtesi del sorgiment d’una nova forma humana, que preludia la modernitat, fa aproximadament un milió d’anys i en algun lloc de l’est d’Àfrica i / o el sud-oest d’Àsia.
La modernitat suposa, entre altres aspectes, un cervell de mida superior als 1.000 centímetres cúbics, un desenvolupament general de l’organisme que s’aproxima ja al nostre, amb infància prolongada i presència del període d’adolescència, desenvolupament dental de tipus modern, proporcions corporals similars a les d’Homo sapiens i una cara que ja recorda a la humana, relativament plana i amb nas prominent. Aquesta modernitat també ve marcada, com és lògic, per la pèrdua dels trets primitius que van definir als hominins anteriors.
Alguns investigadors, com Phillip Rightmire [28], van apostar fa anys per la segregació d’un tipus humà diferent a partir de l’espècie Homo erectus. Aquesta segregació era condició indispensable per explicar l’origen comú dels neandertals i dels humans moderns. L’equip d’Atapuerca va arribar una mica més lluny quan van aparèixer les primeres restes humanes datades en uns 850.000 anys d’antiguitat al jaciment de la cova de la Gran Dolina. Com és ben conegut, l’estudi d’aquests fòssils va culminar el 1997 amb la proposta d’una nova espècie del gènere Homo: Homo antecessor. En llatí, el nom específic significa «explorador», al·ludint així al fet que aquells fòssils trobats a la burgalesa serra d’Atapuerca representaven a les poblacions humanes més antigues conegudes fins la data a Europa. Més tard el 2007 l’equip va trobar una mandíbula al jaciment de la cova de la Sima del Elefante a uns 300 metres de distància de Gran Dolina, la cronologia de la qual es va estimar en 1,2 milions d’anys. I es va classificar com a Homo sp., (és a dir, espècie indeterminada).
La publicació de l’espècie Homo antecessor el 1997 a la revista Science [29] sostenia la hipòtesi que els hominins de la Gran Dolina podien representar a l’ancestre comú dels neandertals i de les poblacions modernes retardant així aquest procés en més de 400.000 anys, hipòtesi formulada per la presència de la cara moderna a ATD6-69 i en altres restes facials d’individus immadurs i adults de TD6. La hipòtesi que els fòssils de TD6 van tenir un paper rellevant en l’evolució humana va ser rebutjada o ignorada per la majoria dels especialistes.
Cap paleoantropòleg està disposat a admetre que unes restes fòssils trobades a l’extrem més occidental d’Euràsia podien estar relacionades amb els humans moderns. Si l’origen d’Homo sapiens es situa a l’Àfrica, com explicar que una espècie europea (Homo antecessor) hagués donat lloc a una espècie africana (Homo sapiens), al mateix temps que originava una espècie genuïnament europea (Homo neanderthalensis)?
L’altra hipòtesi és que l’espècie elegida com a millor candidata per a representar a aquest antecessor comú era Homo heidelbergensis. Aquesta espècie va ser proposada per Otto Schoetensack en 1908, un any després de la troballa de la coneguda mandíbula de Mauer en els sediments sorrencs del riu Neckar i no lluny de la ciutat alemanya de Heidelberg. La seva cronologia ha estat estimada (i acceptada de manera general) en uns 600.000 anys, encara que no ha quedat constància del lloc precís de la troballa.
Mandíbula de Mauer, assignada el 1908 per Otto Schoetensack a l’espècie Homo heildelbergensis.
L’Homo heidelbergensis
Cfr. Tema 20. Homo antecessor, rhodesiensis, heidelbergensis i floresiensis del curs en línia Orígens de l’art i evolució humana: l’homo significans. Pàgs. 6-9. En línia a http://cv.uoc.edu/adf/~04_999_01_u07/llico_20.pdf
Quines eren les característiques d’aquests homínids que per primera vegada es van endinsar en l’Europa profunda? En l’actualitat és possible fer-se una idea molt completa de l’anatomia d’aquests homínids, gràcies a les troballes realitzades al jaciment de la Sima de los Huesos a Atapuerca. Aquesta extraordinària localitat, datada en uns 500.000 anys, ha proporcionat les restes d’una trentena d’individus, corresponents a una població que incloïa mascles, femelles i nens. Algunes d’aquestes restes es troben en un magnífic estat de conservació, com és el cas del crani 5, pràcticament complet, o dels cranis 4 i 6, també molt ben conservats (només els falta la cara). La imatge que ofereixen els homínids de la Sima de los Huesos, així com altres restes de l’entorn europeu assignats a Homo heidelbergensis, és la d’uns individus robustos, que podien arribar fins a 180 cm d’altura i pesar fins a 100 quilos, i que estaven dotats de potents braços i cames. Tant la caixa toràcica com la cintura eren més amples que en nosaltres. En general es tractava d’individus forts i robustos, pitjor dotats per a la marxa que els humans actuals, però probablement més resistents, amb una complexió que anuncia ja la dels neandertals. La seva capacitat craniana se situava al voltant dels 1.200 cc, variant entre els 1.100 cc del crani de Steinheim i els gairebé 1.400 cc del crani 4 de la Sima de los Huesos.
Per la resta, nombroses evidències apunten que els Homo heidelbergensis van ser uns ensinistrats caçadors, les habilitats i domini del medi dels quals superava amb molt la dels homínids de principis del Plistocè. Una de les troballes més espectaculars en aquest sentit correspon al jaciment de Schöningen, a Alemanya. La torbera de Schöningen, datada en uns 400.000 anys, se situa en una mina de carbó prop de Hannover i ha proporcionat abundants restes de grans herbívors. Al costat d’aquesta fauna, de la qual el cavall constitueix l’espècie dominant, van aparèixer també nombrosos útils lítics acheulians. Ara bé, a la tardor de 1996, al costat de diverses restes de cavall, van aparèixer fins a vuit pals allargats de fusta, una de les puntes havia estat acuradament polida per tal de convertir-los en objectes punxants. La longitud mitjana d’aquests objectes, curosament treballats d’un extrem a un altre, és d’uns dos metres, i el seu diàmetre, d’uns quatre centímetres. Es tracta, per tant, d’autèntiques javelines i una de les primeres evidències que els homínids del Plistocè mig disposaven d’una sofisticada tècnica de caça. Així, el fet que les troballes de Shöningen corresponguin a javelines, més que a pesades llances, és altament significatiu. Cal pensar que l’ús de llances, és a dir, de pals punxants que el caçador clavava directament sobre l’animal, va haver d’estar molt més estès del que l’estricta evidència arqueològica deixa entreveure. Així doncs, els homínids del Plistocè mitjà europeu van ser actius caçadors, capaços d’enfrontar-se a grans preses a distància. Un altre element que va haver de jugar un paper determinant en la colonització de les estepes europees va ser el control del foc. En efecte, associat a Homo heidelbergensis, les primeres evidències del seu control es troben en diversos punts de França i Alemanya, totes elles datades al voltant a uns 400.000 anys. Aquest és el cas, per exemple, de Terra Amata, prop de Niça, a la Costa Blava francesa, a on les excavacions dutes a terme per Henri de Lumley van posar al descobert una autèntica llar. Aquest consistia en un forat d’uns 30 centímetres de diàmetre per 15 de profunditat que havia estat envoltat de pedres i al seu interior es van trobar evidències d’argila cremada. Altres llars han estat descrites també a Europa central, per exemple a la localitat hongaresa de Vertesszöllos, a pocs quilòmetres de Budapest, o a Bizingsleben i Shöningen, a Alemanya. També a la conca de Guadix-Basa, la localitat de Solana Zamborino, datada en uns 200.000 anys, va proporcionar evidències de l’ús del foc per part dels homínids del Plistocè mig, a part d’una abundant indústria acheuliana associada a una variada fauna. En aquesta localitat, com a Terra Amata, es va descobrir un cercle de pedres a l’interior del qual van aparèixer restes lítiques i ossos cremats. Aquest control del foc incloïa la capacitat per mantenir-lo i alimentar-lo després d’un incendi fortuït produït per l’acció d’un llamp o qualsevol altre agent natural. Els avantatges del control del foc en les circumstàncies de l’Europa del Plistocè mitjà són evidents. D’una banda, el foc és un element dissuasiu davant de possibles depredadors com els grans felins. A més, molt probablement va constituir un primer element de cohesió del grup durant les fredes nits de l’Europa glacial. El foc proporciona calor i permet una millor cocció dels aliments, millorant la qualitat nutritiva a predigerir les proteïnes i destruint els tòxics de les plantes.
En qualsevol cas, ja fos per l’ús del foc, la caça de grans mamífers, o l’adaptació als freqüents i ràpids canvis climàtics que van caracteritzar a aquest període, els cert és que entre 600.000 i 200.000 anys enrere es va produir un nou augment de la mida cerebral, tant absolut com relatiu, donant lloc a noves capacitats cognitives.
Homo heidelbergensis no és només la primera espècie humana que es va endinsar en les estepes del centre i nord d’Euràsia, sinó també aquella en què detectem les primeres traces d’una mentalitat simbòlica. Així, l’acumulació de restes humanes a la Sima de los Huesos ha estat interpretada com un cas d’inhumació intencional de cadàvers [30]. Per a l’equip d’investigació d’Atapuerca, cap agent natural pot explicar la varietat i el nombre d’individus que es troben junts en aquest extraordinari jaciment. Després de la mort per accident de caça o per malaltia, el grup d’Homo heidelbergensis de la Sima de los Huesos no hauria abandonat els cadàvers dels seus congèneres a la seva sort sinó que els haurien recollit i els hauria abocat dins de la profunda galeria vertical que dóna pas a la cavitat principal del jaciment, evitant així la seva descomposició a l’aire lliure o el seu consum per carronyers. El descobriment en la mateixa Sima d’un únic bifaç bellament treballat i que no hauria estat usat, ha estat interpretat així mateix pels seus descobridors com una ofrena funerària. Si aquesta interpretació és correcta, això indica que fa 500.000 anys aquests homínids tenien ja un sentit de la mort i, probablement, de l’existència d’una ment individual.
L’espècie Homo heidelbergensis va tenir el seu moment de glòria, però molt aviat va passar a formar part del calaix dels records. Les troballes a la Xina van restar protagonisme a la mandíbula de Mauer, encara que durant molts anys va ser sens dubte el fòssil humà més antic del continent europeu. No obstant això, quan científics com Günter Brauer i Chris Stringer van defensar a finals de la dècada de 1980 i principis de la de 1990 la teoria de l’origen africà d’Homo sapiens, es va haver de buscar una espècie que complís amb el requisit desitjat. Alguna mena d’hominí va haver de donar origen als neandertals i als humans moderns. I l’espècie triada va ser precisament Homo heidelbergensis.
Quines van ser les raons per triar aquesta espècie i no una altra? Una combinació d’atzar i de cronologia. La mandíbula de Mauer podia tenir l’antiguitat compatible amb el moment hipotètic postulat pels genetistes (uns 400.000 anys) per a la separació dels avantpassats d’Homo sapiens i d’Homo neanderthalensis. A més, es coneixien en el registre fòssil diversos cranis d’Àfrica (Bodo i Kabwe), Àsia (Dali i Maba) i Europa (Arago i Petralona, i potser altres), que sens dubte s’havien separat de l’espècie Homo erectus. És cert que la cronologia de tots ells segueix sent controvertida, però es poden considerar com a representants de les poblacions del Plistocè Mitjà, potser contemporanis o una mica més recents que la pròpia mandíbula de Mauer. Anatòmicament,
- aquests exemplars tenen un neurocrani gran, capaç d’albergar un cervell d’uns 1.100-1.200 centímetres cúbics, pràcticament d’una grandària molt similar a la de la nostra espècie.
- la forma del cervell d’aquests cranis és primitiva i no difereix de la d’altres avantpassats del gènere Homo, però la seva grandària és clarament superior.
- el crani és més alt i la seva amplada màxima se situa a nivell dels parietals.
- l’os frontal acaba en uns arcs superciliars de molt gruix, però amb forma doblement arquejada i sense formar una visera contínua característica d’Homo erectus.
- l’esquelet facial és gran i sobresortint, amb pòmuls prominents
Els experts van aventurar que tots ells podrien ser inclosos en l’espècie Homo heidelbergensis. La idea que subjau en tot aquest procés té a veure amb la vella hipòtesi d’una continuïtat evolutiva lineal entre les espècies més primitives del gènere Homo i la nostra pròpia espècie. En aquesta successió lineal, tan popular a anuncis i representacions de l’evolució humana, hi hauria formes intermèdies, que s’assemblarien cada vegada més a nosaltres.
Durant els últims 20 anys, les investigacions sobre el genoma d’Homo sapiens i el dels neandertals han permès aproximar-nos d’una manera diferent a la qüestió que planteja el moment de la separació del llinatge d’Homo sapiens i el d’Homo neanderthalensis. S’ha utilitzat sobretot el DNA mitocondrial (mtDNA) de mostres generalment molt nombroses de poblacions humanes vives, així com d’alguns fòssils de neandertals. Entre els especialistes en aquest àmbit destaca Svante Pääbo, una referència en la majoria de les publicacions, sense deixar de citar investigadors com Matthias Meyer, Johannes Krause, Richard Green, Matthias Krings, o Phillip Endicot. Els resultats obtinguts per aquests i altres autors per al moment de la separació del llinatge de la nostra espècie i el llinatge neandertal són molt diversos, i cobreixen un lapse temporal des de fa més de 300.000 anys i 800.000 anys, per assegurar un 95% de confiança en els resultats. Mig milió d’anys representa un període de temps massa ampli com per ser complaents amb aquests resultats. Sens dubte queda molt per fer.
L’equip d’Atapuerca ara ja no proposa que Homo antecessor representi l’últim ancestre comú dels neandertals i de la poblacions modernes, sinó que defensen que aquesta espècie va tenir un paper rellevant en el concepte de modernitat.
Pel que fa al possible escenari convé assenyalar dos fets importants.
En primer lloc, resulta de summa importància destacar l’existència d’un «forat negre» en el registre fòssil dels hominins al voltant del milió d’anys. És molt possible que les condicions per a la conservació de restes orgàniques en aquella època no fossin les més idònies. Repassant la informació disponible, a l’Àfrica podem comptar amb els cranis OH setembre (Etiòpia), Buia (Eritrea) i Daka (Etiòpia), que tenen una antiguitat al voltant del milió d’anys i una capacitat cranial que no arriba o tot just supera els 1.000 centímetres cúbics. A Europa comptem amb una dent de llet d’aproximadament 1,3-1,4 milions d’anys, trobat al jaciment de Barranco León (Granada), i el fragment de mandíbula ATE9-1 del jaciment de la cova de la Sima del Elefante (serra d’Atapuerca), d’1,2 milions d’anys. A més, hi ha greus problemes en la datació dels hominins d’Àsia. En conseqüència, se sap molt poc sobre el que va succeir cap a finals del Plistocè Inferior tant a l’Àfrica com a Euràsia.
En segon lloc, quan es repassa la seqüència biocronològica d’Atapuerca a través del període que cobreix el rang temporal entre 1,3 i 0,7 milions d’anys, els experts són capaços de detectar diversos canvis significatius en la composició de la fauna tant de grans com de petits mamífers. És evident que alguna cosa important va succeir en aquest llarg període, molt probablement relacionat amb els canvis climàtics. A més, l’escàs registre arqueològic d’Atapuerca pertanyent a aquest temps permet inferir un canvi de comportament molt marcat en les poblacions humanes que van viure al voltant del milió d’anys i la població d’Homo antecessor. Els membres d’aquesta espècie van ocupar les coves, i la seva tecnologia, encara que primitiva, comptava amb una estratègia molt definida en la fabricació dels útils. Per contra, les poblacions anteriors eren visitants esporàdics de les coves i la seva tecnologia es preveu molt senzilla. Tot això ens porta a pensar que el sud d’Europa va ser colonitzat una o més vegades per poblacions encara desconegudes fa potser 1,5 milions d’anys. L’espècie Homo antecessor hauria arribat molt després, portant uns costums diferents i una tecnologia més avançada. Es podria, doncs, pensar en una discontinuïtat en el poblament humà d’Europa, amb onades migratòries procedents de regions orientals, que potser es van barrejar entre si a mesura que ocupaven majors extensions territorials.
Una nova hipòtesi: fa aproximadament un milió d’anys es va produir un canvi de ritme en l’evolució del gènere Homo. Els hominins de TD6, amb el seu estrany mosaic de caràcters primitius i derivats, representen l’evidència incontestable d’aquest fet evolutiu. Per descomptat, el canvi va haver de succeir en una única població més o menys nombrosa i en un lloc determinat més o menys ampli. Podem pensar que aquest territori cobria part de l’est d’Àfrica i el sud-oest d’Àsia i es trobava unit a través del Corredor Llevantí, un veritable cordó umbilical durant llargs períodes de temps.
El nostre hipotètic gran centre evolutiu de l’est d’Àfrica i / o del sud-oest d’Àsia va poder ser un punt calent de l’evolució humana, almenys fa una mica més d’un milió d’anys. Els humans de TD6 podrien representar una de les primeres poblacions (si no la primera) que va sortir d’aquest punt calent i va viatjar cap a Europa. Si va ser així, Homo antecessorrepresentaria una branca lateral de la hipotètica població mare que donaria lloc a diferents moments a altres poblacions filles. L’últim avantpassat comú dels neandertals i dels humans moderns s’hauria format en aquest punt calent fa uns 600.000 anys. Experts com Jeffrey Schwartz, Ian Tattersall o Philip Rightmire l’anomenen Homo heidelbergensis. El més important és saber què va passar i com va succeir. El forat negre en el registre fòssil, del qual parlem més amunt, resulta per ara un impediment per identificar i definir el canvi cap a la modernitat. Però tot arribarà.
L’origen dels neandertals
L’anàlisi del mtDNA dels denisovans és molt recent i apunta a l’existència d’un antecessor comú d’aquests humans de les muntanyes Altai de Sibèria, els neandertals i la nostra pròpia espècie en algun lloc indeterminat, però amb una antiguitat d’un milió d’anys
Cap a finals del Plistocè Mitjà, fa uns 250.000 anys, Europa va ser testimoni de l’aparició d’una nova forma de ser humà. Els seus trets facials i dentals ja són presents en els hominins de la Sima de los Huesos d’Atapuerca (430.000 anys d’antiguitat). Els amplis sinus paranasals contribueixen a formar una cara projectada (prognatisme mediofacial) diferent de la nostra. Però el neurocrani dels neandertals és diferent i peculiar. La seva grandària és molt gran i la seva forma, ampla, relativament baixa, allargada i esfèrica en vista posterior, representa una característica exclusiva d’aquests humans. La seva alçada és menor que la dels seus avantpassats del Plistocè Mitjà, amb un cos massís i musculat.
És interessant preguntar-se per l’origen d’aquesta població tan coneguda per tots i explicar el seu èxit evolutiu durant diverses desenes de milers d’anys.
Durant tot el Plistocè Mitjà [31], l’hemisferi nord de la Terra va conèixer fases molt agudes d’empitjorament climàtic. La major part d’Europa va estar coberta per una potent capa de gel durant aproximadament 360.000 anys, repartits en set períodes d’uns 50.000 anys. No podem obviar que els Alps, els Balcans, els Carpats i els Pirineus van representar barreres geogràfiques inexpugnables per als nostres ancestres d’aquesta època. La conseqüència va ser l’aïllament prolongat de nombroses poblacions, gairebé sempre relegades a les penínsules europees, el clima mediterrani de les quals va temperar les condicions glacials i va servir de refugi a plantes i animals. Els humans vam començar a dominar el foc a Europa fa uns 400.000 anys, però aquest transcendental avenç cultural no va ser suficient per viure en regions cobertes per un mantell de gel que va poder arribar a un quilòmetre de gruix. És molt possible que l’aparició dels anomenats neandertals clàssics fos una conseqüència d’aquestes circumstàncies.
Esquema hipotètic de l’evolució del gènere Homo. En aquest esquema es reconeix que Àfrica i Àsia van ser testimonis de l’evolució d’Homo ergaster i Homo erectus, respectivament, en contra dels que postulen que les dues espècies representen un únic taxó. Per a aquests últims, Homo erectus té un recorregut evolutiu d’uns 2 milions d’anys i va viure tant a l’Àfrica com a Euràsia. Per contra, altres investigadors consideren que les diferències morfològiques en el conjunt de fòssils d’aquests dos continents respon no només a l’efecte del temps, sinó a les barreres geogràfiques sostingudes durant milers d’anys. Aquestes barreres i l’«efecte temps» van ser suficients per promoure diferències recognoscibles, que s’ajusten al concepte paleontològic d’espècie.
Els neandertals, però, no van ser una població relegada a un territori concret, sinó que van aprofitar les millores climàtiques dels períodes interglacials per expandir la seva àrea d’influència cap a una gran part d’Euràsia. La presència de neandertals al Corredor Llevantí, a les portes de l’Àfrica, és un bon exemple del seu èxit com a espècie. Si les anàlisis genètiques confirmen la relació dels neandertals amb els denisovans de les muntanyes Altai de Sibèria, serà evident que aquells humans van poder conèixer les poblacions de la Xina i estar a punt d’arribar a les costes de l’oceà Pacífic. Els neandertals van idear una nova forma de tallar la pedra, realitzant eines especialitzades en tasques molt diverses, van controlar el foc mantenint vives les fogueres durant llarguíssims períodes de temps, van ser formidables caçadors socials, van enterrar als seus difunts amb cerimònies simbòliques i patrons definits i alguns indicis apunten cap a un cert coneixement de l’art. L’última prova s’ha localitzat a la cova de Gorham, al penyal de Gibraltar, on els investigadors han descrit gravats senzills en una superfície d’uns 300 centímetres quadrats al terra de la cova. Ningú ha pogut interpretar el significat d’aquests gravats i alguns pensen que representen només un resultat gràfic accidental esdevingut mentre es tallaven trossos de carn o de la pell d’algun animal.
Amb posterioritat a la Sima de los Huesos, fa entre 500.000 i 200.000 anys, les traces d’una certa mentalitat simbòlica es van repetint, encara que sempre embolicades en una considerable polèmica. És el cas d’algunes estatuetes antropomorfes molt tosques, datades cap a 300.000 anys enrere, o de l’ús de pigments fa 280.000 anys. Fa uns 150.000 anys, però, les poblacions europees d’Homo heidelbergensis patiran una transformació profunda, donant lloc a un tipus d’homínid molt característic, Homo neanderthalensis, més conegut popularment com «home de Neandertal», en el qual la evidències d’una mentalitat simbòlica ja són incontrovertibles.
Físicament, els neandertals porten fins a un extrem la tendència a la robustesa corporal que era palesa a Homo heidelbergensis. Els ossos dels seus membres eren més amples i curts que els nostres, i estaven dotats d’unes fortes insercions musculars que denoten un gran desenvolupament de la musculatura. Aquesta robustesa cal interpretar-la en termes de les dures condicions que van haver de resistir els neandertals en el seu hàbitat europeu de l’última glaciació, fa entre 100.000 i 40.000 anys, quan una estepa freda cobria bona part del continent, inclosa la península Ibèrica. El crani dels neandertals és també molt característic, dotat d’uns forts arcs supraciliars que voregen unes grans òrbites arrodonides. La volta craniana era gran i globulosa, i culminava en un engrossiment posterior en forma de «monyo». Els neandertals estaven dotats d’un cervell tan voluminós com el nostre, al voltant de 1.500 cc, tot i que en alguns casos van arribar a superar la mitjana de la nostra espècie. La forma de la cara és també molt característica en els neandertals. Aquesta era molt ampla i prominent, projectada cap endavant (el que es coneix amb el nom de prognatisme), probablement com una conseqüència de la seva robusta dentadura, sobretot pel que fa als seus incisius, molt prominents i adaptats a mastegar i preparar l’aliment abans de la seva ingestió. El prognatisme de la cara del neandertal queda reflectit també en la forma de la mandíbula, que és així mateix molt prògnata. La posició avançada de la dentició a la mandíbula determina en aquest grup l’existència d’un ampli espai entre l’últim molar i la branca ascendent, un caràcter típicament neandertalià. L’orifici nasal era prominent i ample, i també molt alt, el que ha estat interpretat com una adaptació tendent a aïllar la cavitat nasal del fred aire del Plistocè superior.
Potser com a conseqüència de la seva major capacitat cerebral, els neandertals van ser homínids intel·lectualment més avançats que els seus predecessors, autors d’una sofisticada indústria lítica, sobrenomenada «mosterià» o Mode 3. Aquesta indústria incorporava una profunda innovació tecnològica, l’anomenada tècnica Levallois, que permetia l’obtenció de primes làmines tallants i de puntes amb què dotar les llances que utilitzaven durant les seves caceres. Els estris es van fer més petits i diversos, cadascun d’ells especialitzat en una determinada funció, Però és sobretot a nivell de pensament simbòlic on trobem una de les diferències més significatives entre els neandertals i la resta dels seus avantpassats, ja que és amb ells quan apareixen els primers enterraments intencionats. Aquests enterraments s’han trobat al llarg i ample de la seva àrea de distribució, des de La Ferrasie o Le Moustier a França fins Teshik Tash a Uzbekistan, passant per Qafzeh i Kebara a Israel, o Kiik-Koba a Crimea. En aquests enterraments es troben individus de totes les edats, des de nens a gent gran, ja es tracti d’homes o dones. En alguns d’ells hi ha clares evidències que, al costat del cadàver, es van dipositar també objectes valuosos, com eines, cornamentes de bòvids o pigments vegetals.
Així doncs, en els neandertals trobem ja plenament desenvolupat un tipus de comportament que comporta una ment simbòlica, com és el dels enterraments. A part de les implicacions de tipus transcendental que suposa la consciència de la mort, enterrar a un altre congènere significa tenir consciència que ell és com jo, que alguna cosa com el meu «jo» es troba present també en altres individus i que, d’alguna manera , ells són semblants a mi. L’emergència del pensament simbòlic en els neandertals planteja diversos interrogants pel que fa a aquest tipus de comportament. Evidències d’un tipus de pensament simbòlic plenament desenvolupat es troben a l’Àfrica des de fa més de 70.000 anys en els avantpassats africans de la nostra espècie. És molt possible que humans moderns i neandertals coincidíssim a la zona del Pròxim Orient fa uns 80.000 anys i després, més tard, fa entre 30.000 i 40.000 anys a Europa. En quina mesura les manifestacions de pensament simbòlic en els neandertals van poder ser el resultat d’aquests contactes amb els humans moderns? Es tracta d’una possibilitat que no cal descartar. En cas contrari, si els neandertals van desenvolupar les seves pròpies capacitats simbòliques independentment dels avantpassats de la nostra espècie, caldria admetre que aquest tipus de mentalitat hauria aparegut en paral·lel dues vegades en l’evolució humana. Si això és així, i atès que els neandertals posseïen un cervell tan gran com el nostre, caldria admetre que la mentalitat simbòlica és un caràcter lligat a la mida de determinades àrees cerebrals i que aquesta emergeix un cop se supera un determinat Rubicó cerebral.
Tot això s’esdevenia al mateix temps que es gestava l’origen de la nostra espècie a l’altra banda de la Mediterrània.
La tercera humanitat. Homo sapiens
Mentre els neandertals prosperaven a la gelada Europa, a Àfrica l’equivalent local d’Homo heidelbergensis, Homo rhodesiensis, va iniciar també la seva pròpia trajectòria evolutiva, solament que en una direcció molt diferent a la d’aquells. En lloc de primar la tendència a la robustesa, aquells homínids africans van tendir a desenvolupar una anatomia cada vegada més gràcil, gracilitat que va afectar tant a l’aparell locomotor com a les estructures cranials. Així, els arcs superciliares tendeixen a reduir-se, mentre que les òrbites es fan més baixes i quadrangulars. Per sobre d’elles, el front s’alça verticalment. El crani es torna alt i de parets paral·leles. El volum del cervell creix fins a superar els 1.400 cc (cas del crani de Herto, que arriba als 1.450 cc). La cara es fa curta i plana, allotjant un reduït orifici nasal. La reducció dels maxil·lars afecta també la mandíbula, de manera que la dentició es redueix i apareix el mentó, un caràcter fins llavors inèdit en l’evolució humana. Evidències d’aquest nou tipus humà, precursor de la nostra espècie, Homo sapiens, es troben al voltant de fa 200.000 anys en diverses parts d’Àfrica, com Florisbad, a Sud-àfrica i Omo Kibish 2, a Etiòpia. Una mica més recent, entorn a 150.000 anys d’antiguitat, és el crani Djebel Irhoud, al Marroc. No obstant això, la millor representació d’aquest nou tipus d’homínid procedeix de la localitat de Herto, a la vall del riu Awash, a Etiòpia. La mostra inclou un crani adult molt complet, un altre adult fragmentari i un individu infantil, tots ells datats al voltant a uns 160.000 anys. Els fòssils d’Herto (Homo sapiens idaltu) mostren un estadi anatòmic intermedi entre els Homo rhodesiensis del Plistocè mitjà i l’Homo sapiensplenament modern.
Ara bé, el tret més excel·lent d’aquesta nova humanitat rau en la seva mentalitat simbòlica. En una data tan primerenca com fa uns 77 .000 anys, trobem ja a la cova Blombos, a Sud-àfrica, evidències d’aquesta mentalitat en forma d’una tauleta de pigment ocre, en la qual un arcaic habitant de la zona va traçar una sèrie d’aspes , donant lloc a una figura en la qual apareixen triangles i rombes, tallats per línies paral·leles. Quina va ser la intenció de l’autor d’aquells traços? Ens trobem ja davant d’una primitiva expressió d’un sistema de signes, potser relacionats amb el comptatge d’objectes o individus ?, o bé va poder actuar com un primitiu calendari ?, o potser no va existir cap propòsit, com quan algun nosaltres realitza dibuixos sobre un paper mentre parla per telèfon? No ho sabem, però el que sí és innegable és que la tauleta de Blombos denota un nou tipus de mentalitat, un salt qualitatiu en les capacitats simbòliques de la nostra espècie. A la tauleta de Blombos seguirà la producció de tota una nova gamma d’objectes, els quals, a diferència dels útils lítics, són aparentment inútils per a la supervivència, «no serveixen per a res». És el cas de collarets, polseres, arracades o estatuetes.
Tenim evidències que molt aviat aquest nou tipus humà va iniciar una ràpida expansió per Àfrica, expansió que, de nou, el portaria més enllà dels confins d’aquest continent. Així, fa entre 120.000 i 80.000 anys, les restes cranials de Skhul i Qafzeh testimonien que els humans protomoderns estaven ja assentats a Palestina, colze a colze amb les poblacions neandertals del nord. No obstant això, aquesta primerenca colonització del Llevant mediterrani no va desembocar en una veritable sortida de l’Àfrica, ja que fa 50.000 anys de nou trobem els neandertals instal·lats a les localitats d’Amud i Kebara.
Per contra, sabem que cap a aquesta època la nova humanitat moderna va ser capaç d’iniciar una tercera sortida humana d’Àfrica, que el portaria fins a Austràlia, un continent que fins llavors no havia estat trepitjat per cap homínid (i, de fet, per cap primat). Aquestes primeres evidències de presència humana al gran continent-illa estan formades per útils lítics procedents del llac Mungo, dins el districte dels llacs Willandra, al sud-est d’Austràlia, i la seva edat podria arribar als 60.000 anys. Amb posterioritat, en dates que ronden els 60.000 i 40.000 anys enrere, trobem ja restes humanes associades a aquests mateixos útils lítics. Aquestes restes humans, corresponents a un esquelet pràcticament complet i un altre parcial procedents d’un enterrament, mostren una anatomia tant cranial com poscranial gràcil, encara que altres restes humanes posteriors presenten trets més robustos. Els homínids de Mungo presenten trets que els emparenten directament amb els homínids de Herto, d’acord amb l’opinió de Tim White i Berhane Asfaw.
Amb posterioritat, al voltant a 40.000 anys enrere, es produirà la colonització d’Europa per part d’Homo sapiens, evidenciada sobretot per útils lítics corresponent a la cultura aurinyaciana o Mode 4. Les primeres restes humans associats a aquesta cultura són una mica més recents , entorn als 35.000 anys, i corresponen a una mandíbula en bon estat de conservació i altres restes cranials, que inclouen part de la cara, procedents de la cova Pestera cu Oase a Romania. Aquesta ràpida expansió d’Homo sapiens per Europa desembocarà en el progressiu replegament dels neandertals fins a la seva extinció 10.000 anys després.
Fa al voltant de 40.000 anys, es produirà una autèntica explosió cultural, principalment a Europa, que s’ha denominat «la revolució humana». En aquestes dates van tenir lloc importants innovacions, com l’inici del transport i intercanvi de materials a grans distàncies, un important increment en la diversitat morfològica i taxa de canvi dels estris, la fabricació sistemàtica de vestits amb pells, la generalització dels ornaments personals , la sistematització de l’art i les primeres evidències segures de creences i rituals religiosos
. Alguns investigadors, com Richard Klein, atribueixen aquesta revolució cultural a una mutació genètica, la qual s’hauria produït fa uns 50.000 anys, que va millorar la capacitat cognitiva en la nostra espècie. Però cada gen intervé en múltiples funcions, de manera que una sobtada mutació de tal calibre és probable que induís múltiples canvis en l’organització cerebral, els quals tindrien altes probabilitats de resultar desastrosos per a la correcta funció del cervell. És més probable que l’increment de capacitats cognitives i lingüístiques s’hagués anat produint lentament, a través de múltiples mutacions, i que «la revolució humana» fos el resultat d’altres factors, com un increment de la densitat demogràfica o la major necessitat de cooperació entre diferents grups davant els rigors del clima glacial, la qual cosa va poder promoure un major intercanvi de coneixements i idees. Recents troballes mostren que alguns dels canvis culturals associats a la «revolució humana» s’havien donat ja a l’Àfrica fa més de 80.000 anys, el que va en contra de la hipòtesi sostinguda per Richard Klein.
Sigui com sigui, sabem que fa uns 18.000 anys la humanitat moderna va aconseguir finalment trepitjar terra americana. Prèviament, fa uns 40.000 anys, aquests humans s’havien instal·lat a les terres que voregen l’estret de Bering. Potser la colonització d’Amèrica associada al descens de prop de 120 metres del nivell del mar que es va produir durant l’últim màxim glacial i que va convertir l’actual estret de Bering en un istme transitable. Fa uns 16.000 anys tenim evidències de presència humana a Alaska. Des d’allà, van colonitzar la resta d’Amèrica del Nord i és molt possible que iniciessin les primeres incursions a Amèrica del Sud, tal com ho testimonien les eines lítiques del jaciment de Muntanya Verda a Xile, datat en 15.000 anys. En qualsevol cas, fa 11.000 anys Homo sapiens ja s’havia estès de cap a cap en els dos continents americans. Molt probablement gràcies a les seves capacitats simbòliques, la tercera humanitat va aconseguir arribar fins on cap humà modern havia arribat anteriorment, establint la seva presència en els cinc continents.
Lectures de referència
Agustí, Jordi; Bufill, Enric i Mosquera, Marina (2012). El precio de la inteligencia. Barcelona. Crítica. (Col. Drakontos). 256 pàgs.
Allman, John Morgan (2003). El cerebro en evolución. Barcelona. Ariel. (Col. Ariel neurociencia). 1ª impr. 240 pàgs.
Arsuaga, Juan Luis i Martínez, Ignacio (2006). La especie elegida : la larga marcha de la evolución humana. Madrid. Ed. Temas de Hoy. 1ª impr. 448 pàgs.
Briones, Carlos; Fernández, Alberto i Bermúdez, José María (2015). Orígenes. El universo, la vida, los humanos. Barcelona. Crítica. Pàgs. 341-469
Campàs, Joan (2014): 18. Els primers Homo: Homo habilis. Curs en línia: «Els orígens de l’art i l’evolució humana: l’Homo significans». En línia a http://cv.uoc.edu/adf/~04_999_01_u07/llico_18.pdf
Campàs, Joan (2014): 19. El poblament d’Euràsia: l’Homo ergaster/erectus. Curs en línia: «Els Orígens de l’art i evolució humana: l’homo significans». En línia a http://cv.uoc.edu/adf/~04_999_01_u07/llico_19.pdf
Campàs, Joan (2014): 20. Homo antecessor, rhodesiensis, heidelbergensis i floresiensis. Curs en línia: «Els orígens de l’art i evolució humana: l’homo significans». En línia a http://cv.uoc.edu/adf/~04_999_01_u07/llico_20.pdf
Campàs, Joan (2014). 21. L’Homo neanderthalensis. Curs en línia: «Els orígens de l’art i evolució humana: l’homo significans». En línia a http://cv.uoc.edu/adf/~04_999_01_u07/llico_21.pdf
Campàs, Joan (2014). 22. L’Homo sapiens. Curs en línia: «Els orígens de l’art i evolució humana: l’homo significans». En línia a http://cv.uoc.edu/adf/~04_999_01_u07/llico_22.pdf
Carbonell i Roura, Eudal (2005). Homínidos : las primeras ocupaciones de los continentes. Barcelona. Ariel. (Col. Ariel prehistoria ). 864 pàgs.
Cela-Conde, Camilo i Ayala, Francisco (2011). Senderos de la evolución humana. Madrid. Alianza ed. (Col. Alianza ensayo,188). 7ª impr. 632 pàgs.
Leakey, Richard Erskine (2000). El origen de la humanidad. Barcelona. Debate. (Col. Temas de Debate). 1ª impr. 224 pàgs.
Striedter, Georg F. (2005). Principles of brain evolution. Sunderland MA. Sinauer Associates. 436 pàgs.
Tattersall, Ian (2012). Los señores de la tierra. La búsqueda de nuestros orïgenes humanos. Barcelona. Pasado y Presente. 1ª impr. 352 pàgs.
Notes
[1] Leakey, L.S.B.; Tobias, P.V.; Napier, J.R. (1964): «A new species of the genus Homo from Olduvai Gorge». Nature 202, 7-9. En línia a http://www.tarha.ulpgc.es/leakey_1964.pdf
[2] Bernard Wood i Mark Collard: «The changing face of genus Homo». Evolutionary Anthropology, Volume 8, Issue 6, pages 195–207, 1999. En línia a http://www.sfu.ca/~mcollard/documents/Wood_and_Collard_1999b_000.pdf
[3] Semaw, Sileshie i altres (1997). «2,5-million-year-old Stone tools from Gona, Ethipia». Nature 385, 333-336.
[4] Skinner, Matthew i altres (2015). «Human-like hand use in Australopithecus africanus». Science 322, 395-399.
[5] Cadascuna de les petites prolongacions òssies entrecreuades que formen una malla òssia i que limiten, compartimentant, les cavitats medul·lars del teixit esponjós. Espícules òssies en l’os porós que formen una xarxa d’espais que s’intercomuniquen i que estan plens de medul·la òssia
[6] Sonia Harmand i altres: «3.3-million-year-old stone tools from Lomekwi 3, West Turkana, Kenya». Nature 521, 310-315. En línia a http://www.nature.com/nature/journal/v521/n7552/full/nature14464.html
[7] Heinzelin, J. i altres (1999). «Environment and behavior of 2.5-million-year-old Bouri hominids». Science 284, 625-629.
[8] Mohamed Sahnouni i altres: «The first evidence of cut marks and usewear traces from the Plio-Pleistocene locality of El-Kherba (Ain Hanech), Algeria: implications for early hominin subsistence activities circa 1.8 Ma». Journal of Human Evolution 2013 Feb;64(2):137-50
[9] Binford, Lewis Robert (2004). En busca del pasado : descifrando el registro arqueológico. Barcelona. Crítica. 1ª impr. 296 pàgs.
[10] Potts, R., 1998. Early Hominid Activities at Olduvai. Nova York. Aldine de Gruyter. 396 pàgs.
[11] Schrenk, F., Bromage, T., Betzier, C.G., Ring, U., i Juwayeyi, Y.M. (1993). «Oldest Homo and Pliocene biogeography of the Malawi Rift». Nature 365, 833-836. En línia a https://www.researchgate.net/publication/14784975_Oldest_Homo_and_Pliocene_biogeography_of_the_Malawi_Rift_Nature_365_833-836
[12] Asfaw, B., White, T., Lovejoy, O., Latimer, B., Simpson, S i Suwa, G. (1999). «Australopithecus gahri: A new species of Early Hominid from Ethiopia». Science 284, 629-635. En línia a http://www.indiana.edu/~origins/teach/P314/Bouri.pdf
[13] Berger LR, de Ruiter DJ, Churchill SE, Schmid P, Carlson KJ, Dirks PHGM, Kibii JM. (2010)«Australopithecus sediba: A New Species of Homo-Like Australopith from South Africa». Science, 328, 195-204.
[14] Vekua, A., Lordkipanidze, D., Rightmire, G.P., Agustí, J., Ferring, R., Maisuradze, G., Mouskhelishvili, A., Nioradze, M., Ponce de León, M., Tappen, M., Tvalchreridze, M., i Zollikofer, C. (2002). «A new skull of early Homo from Dmanisi, Georgia». Science 297: 85-89.
[15] Lordkipanidze, David, Tea Jashashvili, Abesalom Vekua, Marcia S. Ponce de León, Christoph P. E. Zollikofer, G. Philip Rightmire, Herman Pontzer, Reid Ferring, Oriol Oms, Martha Tappen, Maia Bukhsianidze, Jordi Agusti, Ralf Kahlke, Gocha Kiladze, Bienvenido Martinez-Navarro, Alexander Mouskhelishvili, Medea Nioradze & Lorenzo Rook (2007). «Postcranial evidence from early Homo from Dmanisi, Georgia». Nature 449, 305-310.
[16] David Lordkipanidze, Abesalom Vekua, Reid Ferring, G. Philip Rightmire, Jordi Agusti, Gocha Kiladze, Alexander Mouskhelishvili, Medea Nioradze, Marcia S. Ponce de León, Martha Tappen & Christoph P. E. Zollikofer (2005). «The earliest toothless hominin skull». Nature 434, 717-718.
[17] Lumley, M.A. de i Lordkipanidze, D. (2006). «L’Homme de Dmanissi (Homo georgicus), il y a 1.810.000 ans». Comptes Rendus de l’Académie des Sciences de Paris Paleovol 5, 273-281.
[18] Timothy Bromage i Christopher Dean: «Re-evaluation of the age at death of immature fossil hominids». Nature 317, 525 – 527. 10 October 1985.
[19] Sakai T , Hirata S , Fuwa K , Sugama K , Kusunoki K , Makishima H , Eguchi T , Yamada S , Ogihara N , Takeshita H (2012). « Fetal brain development in chimpanzees versus humans». Current Biology, Volume 22, Issue 18. R791–R792. En línia a http://langint.pri.kyoto-u.ac.jp/ai/en/publication/TomokoSakai/Sakai2012.html
[20] Jeremy de la Silva i Julie Lesnik (2008). «Brain size at birth throughout human evolution: A new method for esimating neonatal brain size in hominins». Journal of HUman Evoluton 55, Issue 6, 1064-1074. En línia a http://www.sciencedirect.com/science/article/pii/S0047248408001401
[21] Swisher, C., Curtis, C.H., Jacob, T., Getty, A.G., Sprijo, A., Widiasmoro (1994). «Age of earliest known homínids in Java, Indonesia». Science 263, 1118-1121
[22] Wanpo, H., Ciochon, R., Yumin, G., Larick, R., Qiren, F., Schwarcz, H., Yonge. C., de Vos, J., Rink, W. (1995). «Early Homo associated with artifacts from Asia». Nature 378, 275-278.
[23] Peter J Obendorf, Charles E Oxnard, Ben J Kefford (2008). «Are the small human-like fossils found on Flores human endemic cretins?». The Royal Society. Biological Sciences. Volume 275, issue 1640. En línia a http://rspb.royalsocietypublishing.org/content/275/1640/1287
[24] Robert B. Eckhardt, Maciej Henneberg, Alex S. Weller, i Kenneth J. Hsü: «Rare events in earth history include the LB1 human skeleton from Flores, Indonesia, as a developmental singularity, not a unique taxon». Proceedings of the National Academy of Sciences of the United States of America. Vol. 111, núm. 33, 11961-11966.
[25] Anita Rauch i altres (2008). «Mutations in the Pericentrin (PCNT) Gene Cause Primordial Dwarfism». Science. 319(5864): 816-9.
[26] Asfaw, B., Gilbert, W.H., Beyene, Y., Hart, W.K. Renne, P.R., Vrba, E.S., White, T. (2002). «Remains of Homo erectus from Bouri, Middle Awash, Ethiopia». Nature, 416, 317-320.
[27] White, T.D. (1985). «Acheulean man in Ethiopia’s middle Awash Valley: the implication of cutmarks on the Bodo cranium». Kroon Memorial Lecture. Netherlands: Joh. Enschede en Zonen Haarlem.
[28] Phillip Rightmire (2008). «Human evolution in the Middle Pleistocene: The role of Homo heidelbergensis». Evolutionary Anthropology 6, 218-227. En línia a http://archeo.uw.edu.pl/zalaczniki/upload23.pdf
[29] Bermúdez de Castro i altres (1997). «A homínid from the Lower Pleistocene of Atapuerca, Spain: possible ancestor to Neandertals and modern humans». Science 276, 1392-1395
[30] Carbonell, E., M. Mosquera, A. Ollé, X.P. Rodríguez, R. Sala, J.M. Vergès, J.L. Arsuaga i J.M. Bermúdez de Castro (2003). «Les premières pratiques funéraires auraient-ils pris place à Atapuerca, il y a 350.000 ans?». L’Anthropologie 107 (1): 1-14.
[31] El Plistocè està datat generalment entre fa 1,8-1,6 milions d’anys i fa 10.000 anys. El nom del Plistocè deriva dels mots grecs pleistos (més) i cenos (nou). El plistocè inferior abasta d’1,8 milions d’anys fins als 780.000 (el paleolític superior abasta dels 2,2 milions d’anys fins els 128.000, i el paleolític mitjà dels 128.000 als 35.000)); el mitjà, fins els 128.000; i el superior fins els 10.000 (el paleolític superior dels 35.000 als 15.000)
Dr. Joan Campàs Aura digital
Estudis d’Arts i Humanitats de la UOC
Curs Som les nostres connexions
Material en format pdf
Tema 0. Presentació i justificació
Tema 1. El cervell ens enganya
Tema 2. El cervell també enganya l’autopercepció del cos
Tema 3. Anatomia cerebral bàsica
Tema 4. Mite: Només utilitzem el 10 % del nostre cervell
Tema 5. Mite: L’homosexualitat (l’hetero, la bi, la trans) és una elecció
Tema 6. Mite: Els fills hereten la intel·ligència dels pares
Tema 7. Mite: Internet i els videojocs són addictius
Tema 8. Mite: El cervell és un ordinador
Tema 9. La memòria
Tema 10. Com definir el «jo»? El cervell i la «joïtat»
Tema 11. L’evolució del cervell (1)
Tema 12. L’evolució del cervell (2)
Tema 13. L’evolució del cervell (3)
Tema 14. L’evolució del cervell (4)
Tema 15. Cervell humà i evolució
Conjunt dels posts amb les introduccions fetes al facebook
Som les nostres connexions