8. El procés evolutiu: Darwin i la selecció natural
27 octubre, 20141. ¿Què podem entendre per evolució?
Evolució s’associa a en Darwin. L’evolució és un procés segons el qual els éssers vius s’han originat els uns dels altres per descendència i canvis. Conseqüència, doncs, de l’evolució és la transformació de les espècies en el curs del temps. .
Tots els animals, plantes i organismes del nostre planeta provenen de formes diferents a partir de les quals han evolucionat.
L’estudi de l’evolució biològica té dos aspectes que es complementen: el procés evolutiu en sí i el dels mecanismes i de les causes de l’evolució. L’estudi del procés evolutiu tracta de la gènesi dels organismes i llurs transformacions successives, i té com a base principal les dades que aporta la paleontologia. Els mecanismes i les causes de l’evolució són objecte d’una branca de la genètica, la genètica evolutiva, que té com a base principal la genètica de poblacions.
El procés evolutiu ve regit per 5 lleis concretes:
1. Llei de Dollo[1] o de la irreversibilitat: quan un animal o membre canvien, no poden tornar a evolucionar al seu estadi anterior.
2. Primera Llei de Cope[2] o de la inespecialització: evolucionen les espècies indiferenciades, les que tenen una més gran capacitat d’adaptació a canvis ecològics.
3. Segona Llei de Cope o de l’increment de la mida: en qualsevol procés evolutiu, les espècies augmenten la seva mida.
4. Llei de Dacqué[3] o del paral·lelisme: en diferents moments de l’evolució, grups diferents d’animals han trobat respostes similars a estímuls similars.
5. Llei de Williston[4]: amb el temps, les espècies tendeixen a simplificar les seves estructures adaptant-se a un medi concret i augmentant amb això el risc de la seva especialització.
2. Mecanismes de l’evolució
L’evolució depèn de diversos processos que tendeixen a modificar la freqüència dels al·lels[5] en el sí de poblacions: mutació, deriva genètica, estrangulació genètica, flux de gens, mescla genètica, selecció natural, selecció sexual, aparellament no aleatori, pedomorfosi[6], pleiotropia[7], pleiotropia antagonista[8]…
L’evolució de les espècies és permesa per les mutacions que experimenten els gens en els cromosomes (constituïts per ADN). Cada ésser viu posseeix ADN (això corrobora la idea d’un origen comú de les espècies): aquesta molècula consta de dues hèlixs complementàries al nivell de les seves bases nitrogenades. Les mutacions poden afectar aquest ADN; són causades per agents mutàgens com els raigs X, alfa, UV, o simplement per una fallada dels orgànuls responsables de la reparació de l’ADN mal transcrit o traduït. Aquestes mutacions afecten la seqüència d’un gen (ordre de les bases de nucleòtids d’un gen: adenina, timina, guanina i citosina).
Aquestes mutacions són la causa del polimorfisme dels gens, és a dir, el fet que dues versions d’un mateix gen (dos al·lels), per exemple, estiguin presents cadascun en un tant per cent de la població de l’espècie considerada. Aquestes mutacions creen, doncs, nous al·lels. Però no cal oblidar el fenomen de les famílies multigèniques, que té un impacte important en l’evolució: un gen pot ser duplicat i traslladat a un altre cromosoma, i tots els gens sorgits d’aquest gen ancestral formen part d’una família multigènica.
L’entorn «enquadra» aquestes mutacions a través d’un procés denominat selecció natural: un gen que presenta un avantatge per a una espècie en un determinat entorn, que permet als seus representants assolir de la millor manera possible la maduresa sexual, s’estén entre els individus de la mateixa espècie, per contra, si és nefast, desapareix. Pel que fa als gens neutres, s’estenen de manera aleatòria, però poden seguir l’evolució (mutacions dels gens homeòtics). Per tant és el medi ambient el que decideix/condiciona l’evolució de les espècies, les quals evolucionen per estar sempre més adaptades a l’entorn. Per tant, el caràcter aleatori de les mutacions de l’ADN es veu compensat per la selecció ambiental.
És molt important no confondre evolució i innovació; cal tenir en compte que l’espècie que sobreviu és l’espècie millor adaptada, durant un cert període, al seu entorn.
A més, és important no veure l’evolució des d’un punt de vista genealògic, sinó filogenètic, atès que les espècies no descendeixen les unes de les altres. Fenòmens com la deriva genètica provoquen que dues poblacions d’una mateixa espècie aïllades durant un període molt llarg de temps divergeixin i formin dues noves espècies. Per exemple, per al llinatge humà, l’arbre filogenètic és ramificat: diverses espècies Homo i Australopithecushan viscut simultàniament. Cal subratllar que els humans –contràriament a la creença popular– no descendeix del «mico», sinó que tenim un avantpassat en comú amb ell. I aquest ancestre comú no era un mico. El nostre patrimoni genètic és molt similar al dels ximpanzés, en realitat les diferències entre nosaltres i ell es redueixen a la diferència d’uns pocs gens crucials: ens permeten mantenir caràcters juvenils durant tota la nostra vida, que ens permet mantenir el bipedisme (el petit ximpanzé és gairebé bípede) i allarguen la fase embrionària, que permet la mitosi de moltes més neurones.
Parlem de filogènesi, no de genealogia. No es tracta d’un procés lineal, ni de la desaparició d’una espècie donant lloc a una altra. Ex. Australopithecus, Homo habilis i erectus varen coexistir en el temps.
L’evolució resulta de l’acció de diversos mecanismes evolutius susceptibles d’alterar la freqüència dels genotips[9] en una població, i per tant modificar caràcters morfològics, fisiològics o de comportament.
El procés és en tots els casos idèntic, en el mateix ordre:
- aparició d’un nou caràcter hereditari en un o uns pocs individus.
- aquest nou caràcter s’estén per les poblacions, ja sigui perquè és més adaptat al medi, ja sigui per un efecte de l’atzar.
A cada generació, noves variants genètiques apareixen que poden transmetre el seu patrimoni genètic en el moment de la reproducció. Aquestes fonts de diversitat genètica són les següents:
- mutacions puntuals en la seqüència d’ADN dels gens;
- reordenaments cromosòmics;
- recombinacions o intercanvis genètics que es produeixen durant la reproducció sexual i, en els bacteris, en la transferència d’ADN entre les cèl·lules.
Podem afegir a aquesta llista una altra font de diversitat genètica com són les migracions mitjançant les quals l’estoc genètic en una població donada es veu renovat per l’arribada d’altres membres de l’espècie portadors d’un patrimoni genètic diferent.
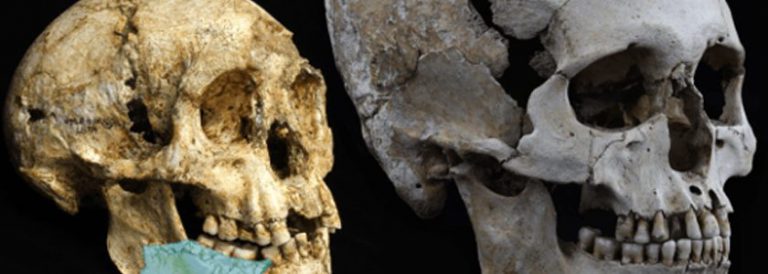
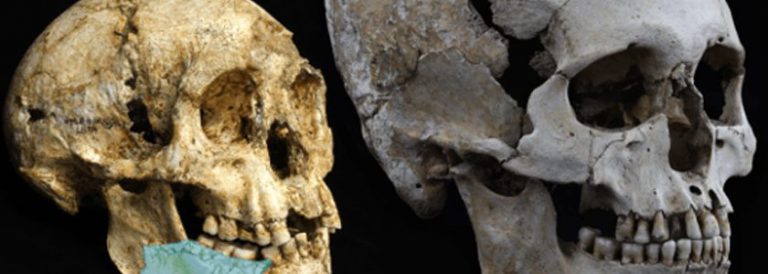
Recents estudis que han permès reconstruir l’ADN d’un os humà datat en 400.000 anys d’antiguitat. Fèmur humà d’on s’ha extret l’ADN. Els habitants d’Atapuerca sembla que no són els ancestres dels neandertals, i estan emparentats amb els humans que visqueren a Denísova (Sibèria) fa 40.000 anys. La investigació indica que els humans de la Sima de los Huesos es varen separar del llinatge dels neandertals i dels Homo sapiens fa un milió d’anys. La família humana es va dividir llavors en dues branques: una va evolucionar cap els humans de la sima i de Denísova, que es varen separar fa 730.000 anys; l’altra, cap els Homo sapiens i els neandertals, que es varen separar fa 500.000 anys.
3. La mutació
Les mutacions[10] són el resultat d’errors durant la replicació dels gens. Aquestes modificacions accidentals poden ser transmeses a la descendència si afecten les cèl·lules germinals, donant lloc a individus portadors de nous gens, que cap dels seus parents manifestaria.
Hi ha diversos tipus de mutacions (mutacions puntuals, duplicacions de gens, ruptures cromosòmiques, insercions o delecions de seqüències). Aquestes diferents modificacions són suficients per explicar la diversitat genètica observada en la natura.
S’hi poden afegir les mutacions per intercanvi de material genètic entre espècies, diferents mecanismes naturals:
- els bacteris són capaços d’integrar i utilitzar el material genètic (plasmidis) intercanviat entre dos bacteris, o simplement presents en el medi. En la majoria dels casos això no condueix immediatament a la definició d’una nova espècie, però el fenomen pot contribuir i contribueix de manera significativa a la selecció natural (per exemple, nombroses resistències als antibiòtics es propaguen d’aquesta manera).
- els virus poden servir com a vectors per transferir material genètic d’una espècie a una altra, fins i tot molt diferent; en la majoria dels casos, la infecció viral provoca la mort de la cèl·lula víctima, però no necessàriament , i la cèl·lula diana pot participar en la reproducció (especialment en les plantes). Un dels exemples més coneguts (i àmpliament utilitzat en la investigació) és el del virus del mosaic del tabac.
- les plantes es poden hibridar, donant lloc a noves espècies (el blat n’és un exemple típic).
- finalment, cal esmentar les simbiosis estrictes, que són creacions de noves espècies a partir d’antigues, però sense intercanvi ni fusió del material genètic (almenys inicialment). Líquens i, segons la hipòtesi de l’endosimbiosi, els orgànuls anomenats «autònoms» a l’interior de les cèl·lules (cloroplasts, mitocondris) són d’aquest tipus d’associació.
Les mutacions augmenten la variabilitat genètica dins d’una població. No obstant això, poques vegades són beneficioses per a l’individu que les porta atès que poden destruir l’activitat d’un gen essencial.
4. Models interpretatius del segle XIX
La teoria de Lamarck: ¿La necessitat crea l’òrgan?
Malgrat les aportacions de Bufont, Kant i Laplace, la de Lamarck (1744-1829)[11] és la primera formulació de la teoria de l’evolució, que va exposar a l’obra “Philosophie zoologique” (1809).
Dues són les seves idees bàsiques: la necessitat crea l’òrgan i la fixació hereditària dels canvis aconseguits.
Segons Lamarck, la naturalesa ha produït, per generació espontània, els éssers vius més simples, i tots els altres s’han format successivament a partir d’ells, fins arribar als d’organització més complexa. Les influències del medi extern actuen modificant les necessitats de l’animal i aquest elabora una resposta primària adaptant-se a les noves circumstàncies.
L’evolució, doncs, és una progressió de la naturalesa. L’ésser viu s’adapta al seu entorn per l’adquisició de caràcters que transmet als seus descendents.
Així, si a causa d’un canvi en el medi extern, un animal no ha de fer servir un dels seus òrgans, aquest es reduirà i s’atrofiarà, fins arribar, fins i tot, a desaparèixer; i, contràriament, la utilització d’un òrgan comporta el seu enfortiment i, fins i tot, la creació d’un nou òrgan, com a resposta a les necessitats. Aquestes variacions són transmeses per l’individu als seus descendents.
Aquesta teoria es resumeix en dues lleis:
- L’ús més freqüent d’un òrgan el fortifica, el desenvolupa i li confereix una potència proporcionada a la durada d’aquest ús; la manca constant d’ús d’un òrgan l’afebleix, en fa minvar progressivament les facultats i, fins i tot, acaba fent-lo desaparèixer.
- Tot allò que la naturalesa ha fet adquirir o perdre als individus, ho conserva a través de la generació en els nous individus, sempre que els canvis siguin comuns als dos sexes.
Fou el primer a afirmar la relació existent entre tots els éssers vius i llur comunitat d’origen, enfront de les idees fixistes i creacionistes postulades pels seus predecessors.
Les hipòtesis de Darwin: ¿L’èxit de l’espècie depèn de la lluita de l’individu per a la supervivència?
Les observacions de Darwin (1809-1882)[12] sobre la distribució geogràfica dels fòssils de l’est de l’Amèrica del Sud i la fauna de les illes Galápagos (arran del seu viatge amb el Beagle) el convenceren que els diferents animals de la natura no podien ser explicats per creació separada, sinó que llur origen demanava una teoria evolucionista.
En el seu llibre L’origen de les espècies (1859) presenta el cas de l’evolució general i desenvolupa la teoria de la *selecció natural.
Bé que es dóna la tendència de tots els organismes a augmentar en una progressió geomètrica, hom pot comprovar que el nombre d’individus d’una espècie determinada resta més o menys constant. D’aquí dedueix que hi ha una lluita per l’existència, una lluita universal per la conservació de la vida, i que de la lluita de l’individu per la supervivència depèn l’èxit de la perpetuació de la progènie. La lluita per l’existència no és una lluita cruenta, sinó una millor adaptació a l’ambient.
La idea central és, doncs, que els éssers vius més *adaptats al medi ambient són els que sobreviuen, mentre que els menys adaptats són eliminats.
Entre els animals de la mateixa espècie aquesta lluita es planteja, sobretot, en el camp de la recerca de l’aliment, en el de la fugida dels enemics, en la resistència enfront de les malalties i en el de l’èxit en l’aparellament.. Tenint en compte que existeixen variacions entre els individus de la mateixa espècie i que en neixen més que no en sobreviuen, tindran més oportunitats de perpetuar l’espècie aquells individus que tinguin algun avantatge i seran destruïts aquells que presentin variacions en sentit contrari. Aquest mecanisme, la selecció natural, explica l’origen de les espècies.
Mitjançant l’acumulació successiva de les variacions en una mateixa direcció, es podria produir finalment una nova espècie.
Quin fou el procés evolutiu que, segons Darwin, va separar els éssers humans dels altres primats?
Darwin argumentà que l’esquelet -excepte el crani- va adquirir les seves proporcions molt abans que el cervell i el cap assolissin les característiques típicament humanes. Això mostra que la selecció natural va incidir en la capacitat humana d’adaptar-se culturalment i socialment a un medi ambient ple d’espècies més fortes físicament, i reafirma, així, la idea que els humans som un animal cultural.
Subratllà les condicions prèvies materials necessàries per a l’aparició de la societat humana. Esquemàticament: amb el bipedisme, les mans han estat alliberades de la locomoció, cosa que permet, per una banda, la fabricació d’instruments i, per l’altra la utilització de les mans per a la defensa; com a conseqüència d’aquest fet, es redueixen les dents i la mandíbula, ja que no s’han de fer servir per a l’atac o la defensa, i s’incrementa el volum cranial, amb el que s’aconsegueix un increment de l’enginy i la destresa que, al seu torn, repercutirà en la fabricació d’estris. Tot plegat desemboca en l’èxit en la competència amb d’altres animals.
Considerà que els avantatges adaptatius de l’espècie humana exigiren un gran desenvolupament del que anomenà instints socials. La cooperació comunitària en l’obtenció d’aliments i les accions defensives i ofensives, afegida a l’augment d’intel·ligència, està a la base de la responsabilitat social i fonamenta la moral humana. La moral, doncs, va donar avantatges a les primeres comunitats humanes en la seva adaptació mitjançant un augment de la cooperació interna en el grup.
Va partir del supòsit que l’origen de la moral era doble:
- l’instint de simpatia: creu que una mena d’esperit de cooperació és un tret innat pels avantatges adaptatius que suposa; a la llarga s’obtenen beneficis de la col·laboració i la solidaritat.
- la regulació de l’activitat sexual: creu que les unions heterosexuals estables foren essencials per al desenvolupament de les primeres societats humanes. El matrimoni fou una solució cultural a la gelosia sexual dels mascles.
Vinculà, sistemàticament, els conceptes d’home-organisme-en-evolució, amb el d’home-fabricant-d’estris i el d’home-caçador, i així ens presenta el comportament masculí i femení: els homes tenen més impulsos sexuals i són més agressius i competidors. Les dones no són agressives i tendeixen a tenir cura dels fills. La seva naturalesa asexuada explica el seu important paper en l’evolució, consistent a frenar i controlar l’energia del mascle, i així estimula la cooperació a l’interior del grup perquè redueix les tensions sexuals.
Darwin i la selecció natural
En biologia, la selecció natural és un dels mecanismes que causa l’evolució de les espècies. És un mecanisme particularment important perquè explica l’adaptació de les espècies al seu entorn al llarg de generacions. La teoria de la selecció natural permet explicar i entendre com l’entorn influeix en l’evolució de les espècies i les poblacions seleccionant els individus més adaptats.
La seva idea central és el fet que els trets que afavoreixen la supervivència i la reproducció incrementen la seva freqüència d’una generació a una altra. Això resulta del fet que els portadors d’aquests trets tenen més descendents i aquests darrers tenen també aquests trets perquè són hereditaris.
«He donat a aquest principi, en virtut del qual una variació per insignificant que sigui es conserva i perpetua, si és útil, el nom de selecció natural».
El terme «selecció natural» fou imaginat per Darwin per analogia amb la selecció artificial practicada pels humans des de fa mil·lennis: els pagesos o pastors elegeixen de cada collita o generació d’animals els individus que presenten les millors característiques per fer-les reproduir-se. El mecanisme de selecció darwiniana permet explicar de manera naturalista la complexitat adaptativa dels éssers vius, sense haver de recórrer a la intervenció sobrenatural o d¡origen diví.
Aquesta teoria, tal com va ser inicialment descrita per Darwin, descansa en tres principis: el principi de variació, el principi d’adaptació i el principi d’herència.
En general, en les poblacions d’individus d’una mateix espècie, existeixen diferències més o menys importants entre aquests individus. En biologia, s’anomena caràcter, tot allò que és visible i que pot variar d’un individu a un altre. Hom diu que existeixen diferents tretsper un mateix caràcter. Per exemple, en l’ésser humà, el color e la pell o dels ulls són caràcters per als quals existeixen diverses variacions o trets. La variació d’un caràcter en un individu donat constitueix el seu fenotip. Aquesta és la primera condició per tal que hi hagi selecció natural: al sí d’una població, alguns caràcters han de presentar variacions: és el principi de variació.
Alguns individus tenen variacions que els permeten reproduir-se més que als latres, en un entorn concret. Disposaran llavors d’un avantatge selectiu sobre els seus congèneres:
- la primera possibilitat és, per exemple, que en escapar millor als depredadors, , en agafar menys malalties, en accedir més fàcilment als aliments, aquests individus arriben més fàcilment a l’edat adulta per a ser aptes per a la reproducció. Aquells que tenen una millor capacitat de supervivència podran, doncs, reproduir-se més.
- en el cas particular de la reproducció sexual, els individus que hagin sobreviscut poden ser portadors d’un caràcter particularment atractiu per als companys del sexe oposat. Aquests seran capaços d’engendrar una més gran descendència pel fet de copular més.
En els dos casos, l’increment de la capacitat de sobreviure i de reproduir-se es tradueix en un augment de les taxes de reproducció i, per tant, d’una descendència més nombrosa, per als individus portadors d’aquestes característiques. Es diu llavors que aquest tret del caràcter donat ofereix un avantatge selectiu, en relació als altres. És en aquest principi d’adaptació únicament on intervé el medi.
La tercera condició per a que hi hagi selecció natural és que les característiques dels individus han de ser hereditàries, és a dir, que es puguin transmetre a la seva descendència. Alguns caràcters, com el bronzejat o saber-se les capitals dels estats africans, no depenen del genotip, és a dir del conjunt dels gens de l’individu. En la reproducció, són els gens que, transmesos als descendents, comportaran el pas de certs caràcters d’una generació a una altra. És el principi d’herència.
Aquests tres primers principis provoquen, doncs, que les variacions hereditàries que confereixen un avantatge selectiu seran més transmeses a la següent generació que les variacions més avantatjoses. Efectivament, els individus que són portadors de les variacions avantatjoses es reprodueixen més. Al llarg de les generacions es podrà veure la freqüència dels gens desavantatjosos disminuir fins arribar, fins i tot, a desaparèixer, mentre que les variacions avantatjoses s’estendran per la població, fins arribar, eventualment, a ser compartides per tots els membres de la població o de l’espècie. Per exemple, el bipedisme és un caràcter comú a tots els éssers humans moderns.
Recordem la divertida història[13] atribuïda a Richard Dawkins. Dos brontosaures veuen un tiranosaure rex avançar cap a ells i apreten a córrer tan com poden. Al cap d’una estona un diu a l’altre: «Per què ens cansem tant? No tenim la més mínima oportunitat de córrer més de pressa que un tiranosaure!» I l’altre li respon amb cinisme: «Jo no intento córrer més de pressa que el tiranosaure. Intento córrer més de pressa que tu!». La idea és recordar que el procés té menys a veure a una competència entre espècies, que a una competència a l’interior de cada espècie.
Quan observem espècies en el seu entorn, semblen estar totalment adaptades al seu medi: el llarg coll i potes de les girafes estan ben adaptats per arribar a les fulles altres de les acàcies. Es podria dir que són els organismes no adaptats els qui no han sobreviscut en aquest medi.
A més, algunes variacions avantatjoses en un entorn donat poden esdevenir nefastes en altres condicions. Per exemple, en un entorn cobert de neu, una pell o pelatge blancs permet no ser vist per la presa o pel seu depredador, però si el medi esdevé boscós o més fosc perdrà el seu avantatge selectiu. La conseqüència d’aquest fenomen és, doncs, que al llarg de les generacions, per la selecció natural, els caràcters observats en una població estaran més o menys adaptats a les evolucions del seu ecosistema.
Forma típica i forma carbonaria, d’una arna que està sobre el mateix arbre. La forma típica és més difícil de veure en un arbre dins una zona sense contaminació i es camufla millor dels seus depredadors.[14]
Els factors de l’entorn que poden, doncs, provocar una selecció natural poden ser:
- factors fisicoquímics (el biòtop): clima, medi terrestre, aquàtic…
- altres éssers vius (la biocenosi): presència de depredadors, paràsits, microbis, competidors…
L’adaptació de les espècies al seu nínxol ecològic pot, de vegades, portar a dues espècies que ocupen un medi similar, a adquirir similituds que no són degudes al seu eventual parentiu. Es parla, en aquest cas, d’evolució convergent. Aquest fenomen s’interpreta com el fet que les mateixes pressions del medi porten a les mateixes solucions adaptatives. Els ulls dels vertebrats i dels cefalòpodes constitueixen un dels exemples més clars de convergència atès que l’avantpassat comú d’aquests dos taxons no posseïa ulls complexos.
Aquests casos de convergència evolutiva són sovint posats de manifest per argumentar a favor d’una concepció adaptacionista de l’evolució per selecció natrual segons la qual l’essencial de les característiques observades en les espècies vives no són degudes a l’atzar sinó que són el resultat de diverses pressions al llarg de la història evolutiva de les espècies.
La capacitat de l’adult de digerir la lactosa de la llet ha estat seleccionada fa uns 9.000 anys en les poblacions humanes originàries del nord d’Europa o de l’Àsia central on es dedicaven al pasturatge del bestiar per la seva llet, però no en altres poblacions on aquest aliment és absent[15]. Recentment, s’ha descobert que aquesta capacitat era present igualment en algunes ètnies e l’est d’Àfrica, els Tutsis i Peuls del Sudan, de Tanzània i de Kènia, però a partir d’altres tres mutacions, seleccionades independentment l’una de l’altra[16]. Aquestes haurien aparegut fa entre 7.000 i 3.000 anys. Es tracta d’un exemple de convergència evolutiva.
La persistència d’una malaltia genètica com l’anèmia de cèl·lules falciformes, o anèmia drepanocítica és deguda al fet que l’al·lel responsable de la malaltia confereix també un avantatge selectiu en augmentar la resistència al paràsit responsable del paludisme.
5. Models interpretatius del segle XX
La teoria sintètica de l’evolució constitueix el marc conceptual més utilitzat en l’estudi científic dels processos evolutius de la biologia. Aquesta teoria es basa en la integració de la teoria de l’herència mendeliana i de la genètica de les poblacions a la teoria de Darwin. Aquesta síntesi es va dur a terme en els anys 1930 i 1940 per R.A. Fisher, J.B.S. Haldane, Sewall Wright, Julian Huxley, Ernst Mayr, Bernhard Rensch, George Gaylord Simpson i George Ledyard Stebbins. Anomenada així per Julian Huxley el 1942, també es coneix aquesta teoria com a neodarwinisme o síntesi darwiniana per posar en relleu el fet que es tracta d’una extensió de la teoria original de Charles Darwin que no coneixia els mecanismes de l’herència genètica.
En les primeres dècades del segle XX, la majoria dels naturalistes seguien creient que el lamarckisme i l’ortogènesi[17] eren la millor explicació de la complexitat del món, però el desenvolupament de la genètica va convertir aquesta posició en insostenible. Theodosius Dobzhansky, un post-doctorant que treballava al laboratori de T. H. Morgan havia estat influenciat pels treballs sobre la diversitat genètica de Sergei Chetverikov. Ell va ajudar a tancar la bretxa entre la microevolució desenvolupada pels genetistes de les poblacions i la macroevolució observada pels biòlegs sobre el terreny amb el seu llibre Genetics and the Origin of Species[18] aparegut el 1937. Va estudiar la diversitat genètica de les poblacions «primitives» i va demostrar que, contràriament a les suposicions dels genetistes de les poblacions, aquestes poblacions presentaven una gran varietat genètica amb diferències marcades entre les subespècies. El llibre també transcriu els treballs matemàtics dels genetistes a un llenguatge més entenedor. A Gran Bretanya, Edmund Brisco Ford, el pioner de la genètica ambiental, va seguir treballant durant els anys 1930-1940 per demostrar el poder de la selecció natural, a través de factors ecològics, que incloïa el manteniment de la diversitat genètica gràcies al polimorfisme com els grups sanguinis en els éssers humans.
El biòleg Ernst Mayr fou influenciat pels treballs del biòleg alemany Bernhard Rensch mostrant la influència de factors ambientals locals sobre la distribució geogràfica de subespècies i espècies estretament relacionades. Mayr segueix els treballs de Dobzhansky, i en el seu llibre de 1942 Systematics and the Origin of Species[19], emfasitzava el paper del caràcter vicariant en l’aparició de noves espècies. Mayr també va formalitzar el terme espècie com «grups de poblacions naturals, efectivament o potencialment capaços d’interfecundar-se, que estan genèticament aïllades d’altres grups similars».
En el seu llibre de 1944, Tempo and Mode in Evolution[20], George Gaylord Simpson va mostrar que els registres fòssils eren consistents amb el patró aleatori i irregular predit per la teoria sintètica de l’evolució i que la descendència lineal que mostrava la realitat del lamarckisme i l’ortogènesi era només una il·lusió. El 1950, George Ledyard Stebbins va publicar Variation and Evolution in Plants que permet la integració de la botànica en la teoria sintètica. L’aparició d’un consens interdisciplinari en els treballs sobre l’evolució serà anomenat la teoria sintètica de l’evolució seguint el llibre de Julian Huxley (nét de Thomas Henry Huxley) Evolution: The Modern Synthesis[21].
La teoria sintètica de l’evolució proporciona un marc sòlid per a unir la teoria darwiniana amb la genètica mendeliana. Va ajudar a consolidar la legitimitat de la biologia de l’evolució. La síntesi també va comportar igualment una contracció significativa del pensament evolutiu (el que Stephen Jay Gould denomina «enduriment de la síntesi»). Des de la dècada de 1950, la selecció natural es va convertir en l’únic mecanisme acceptable per explicar els canvis evolutius i la macroevolució era considerada simplement com el resultat de la microevolució.
A mitjan segle XX va començar el sorgiment de la biologia molecular i amb ella la comprensió de la naturalesa dels gens com a seqüències d’ADN i les seves relacions a través del codi genètic fins a la fabricació de proteïnes. Al mateix temps, la millora de les tècniques d’anàlisi de proteïnes introduïren els processos bioquímics en la teoria sintètica de l’evolució. A principis de la dècada de 1960, els bioquímics Emile Zuckerkandl i Linus Pauling varen proposar la hipòtesi del rellotge biològic que estipulava que les mutacions genètiques s’acumulen a un ritme constant al llarg del temps, cosa que permetia calcular el punt en el qual dues espècies s’havien separat. A partir de 1969, Motoo Kimura[22] i altres van proposar una base teòrica per al rellotge biològic, avançant que, almenys en el nivell molecular, la majoria de les mutacions genètiques no comportaven ni un avantatge ni un desavantatge i que la deriva genètica (més que la selecció natural) comportava la majoria dels canvis genètics: la teoria neutralista de l’evolució. A la dècada de 1960, la biologia molecular era vista com una amenaça per al centre tradicional de la biologia evolutiva. Els arquitectes de la teoria sintètica de l’evolució eren molt escèptics envers l’aproximació molecular en especial quan es connectava a la selecció natural.
A mitjan 1960, George C. Williams va criticar fortament les explicacions de l’adaptació basades en el terme de «supervivència de l’espècie» (argument de la selecció de grup). Aquesta visió fou en gran mesura reemplaçada per la teoria del gen egoista, encarnada per la selecció de parentiu promoguda per William Donald Hamilton, George R. Price i John Maynard Smith. Aquesta teoria es resumeix i popularitza en el llibre The Selfish Gene(El Gen Egoista), de Richard Dawkins[23].
El 1973, Leigh Van Valen va proposar el terme «Reina Roja», tret del llibre A l’altra banda del mirall de Lewis Carroll per descriure la situació de “cursa armamentista” evolutiva duta a terme per les espècies que han d’evolucionar constantment simplement per seguir el ritme imposat per les espècies amb les quals estan en coevolució. Hamilton, Williams i altres suggerien que aquesta idea podia explicar l’evolució de la reproducció sexuada: l’augment de la diversitat genètica permesa pel sexe permet mantenir una resistència contra les espècies competidores, els paràsits i els factors ambientals tot i l’alt cost d’aquesta manera de reproducció.
Actualment, l’evolució ja no és vista com la transformació d’individus aïllats, sinó com la de grups d’individus de la mateixa espècie, és a dir, de poblacions. Però el principi bàsic és el mateix: s’explica l’evolució per l’acció de la selecció natural sobre les poblacions.
Una població evoluciona quan la freqüència d’una versió d’un gen anomenat al·lel (o de diversos al·lels) s’hi modifica. Es veu així estendre’s en algunes espècies caràcters que han adquirit, a causa dels canvis ambientals, un valor adaptatiu que no posseïen abans; els individus portadors d’aquests caràcters són especialment afavorits en el nou entorn, en el qual s’hi trobaven en certa mesura «preadaptats»; constitueixen llavors ràpidament una gran part de la població o fins i tot tota la població de l’espècie. Els caràcters que s’expandeixen corresponen als al·lels que ja existien «silenciosament» al sí de l’espècie.
Quan el conjunt dels individus que constitueixen una espècie forma diverses poblacions aïllades, cadascuna d’aquestes poblacions pot adquirir característiques particulars i donar lloc a diferents varietats dins de la mateixa espècie. Si aquestes variacions són, després, incapaces de creuar-se, divergeixen cada cop més i finalment són interestèrils: constitueixen, llavors, espècies diferents.
La teoria sintètica és considerada el model explicatiu més sòlid dels que es disposa actualment per comprendre els fenòmens evolutius. Tanmateix, fins i tot subratllant que realment no existeix avui una sòlida teoria alternativa desenvolupada, alguns científics sí han reclamat la necessitat de realitzar una reforma o ampliació de la Teoria Sintètica, amb nous models capaços d’integrar la biologia del desenvolupament o incorporar una sèrie de descobriments biològics el paper evolutiu dels quals s’està debatent, tals com certs mecanismes hereditaris epigenètics, la transmissió horitzontal; o propostes com l’existència de múltiples nivells jeràrquics de selecció o la plausibilitat de fenòmens d’assimilació genòmica per explicar processos macroevolutius (increment de complexitat per integració com complement a l’increment en complexitat per transformació –gradual–).
Lynn Margulis va més enllà de la seva teoria científica de la simbiogènesi, per postular una nova teoria en la qual afirma que la simbiosi seria la font principal de la variació heretada, mitjançant la qual es combinen genomes senceres. Tanmateix a diferència de la seva acceptada i provada teoria sobre l’origen de les cèl·lules eucariotes, la nova teoria de Lynn Margulis sobre la simbiosi entre microorganismes com a important força de l’evolució, no és acceptada per la comunitat científica, per manca d’evidència contundent (no explicable per les hipòtesis vigents) a favor.
[1] Dollo, Louis(1893) : «Les lois de l’évolution». Bulletin de la Société Belge de Géologie de Paléontologie & d’ Hydrologie, Volum VII: pp. 164-166. En línia a http://www.paleoglot.net/files/Dollo_93.pdf
[2] Cope, Edward Drinke.: «On the Evolution of the Vertebrata, Progressive and Retrogressive». The American Naturalist Volum19, nº 3, març 1885: 140–148. En línia a http://www.jstor.org/stable/2450075
[3] Dacqué, Edgard (1928). Das fossile Lebewesen: Eine Einführung in die Versteinerungskunde. Berlín. Springer. 184 pàgs.
[4] Williston, Samuel Wendall (1914). Water Reptiles of the Past and Present. Chicago: University of Chicago Press.
[5] S’anomenen al·lels a les diferents versions d’un gen. Cada al·lel difereix en una o més diferències de la seqüència de nucleòtids. Aquestes diferències sorgeixen per mutació en la història de l’espècie, o per recombinació genètica. Tots els al·lels d’un gen ocupen el mateix locus (emplaçament) en el mateix cromosoma.
[6] És la conservació del caràcter larvari en l’adult reproductor
[7] Del grec pleio, molts, i tropo, canvis) és el fenomen pel qual un sol gen és responsable d’efectes fenotípics o caràcters distintius i no relacionats.
[8]És l’expressió de variants genètiques (al·lels d’un gen) que afecten diversos caràcters, de manera favorable en les primeres etapes de la vida i de manera desfavorable en les posteriors. Aquesta formulació és una de les que, des de l’evolucionisme, ha permès explicar en termes genètico-poblacionals la fortalesa de les etapes reproductives i el declivi físic de la vellesa.
[9] El concepte de selecció natural és anterior a la comprensió de la genètica, el mecanisme de l’herència per a totes les formes de vida conegudes. En termes moderns, la selecció actua sobre el fenotip d’un organisme, o característiques observables, però és la genètica de l’organisme o genotip el que s’hereta. Aquest és l’enllaç entre la selecció natural i la genètica.
[10] Les mutacions són la matèria primera de l’evolució. L’evolució té lloc quan una nova versió d’un gen, que originalment sorgeix per una mutació, augmenta la seva freqüència i s’estén a l’espècie gràcies a la selecció natural o tendències genètiques aleatòries (fluctuacions casuals en la freqüència dels gens). Abans es pensava que les mutacions dirigien l’evolució, però en l’actualitat es creu que la principal força directora de l’evolució és la selecció natural, no les mutacions. No obstant això, sense mutacions les espècies no evolucionarien.
La selecció natural actua per incrementar la freqüència de les mutacions avantatjoses, que és com es produeix el canvi evolutiu, ja que aquests organismes amb mutacions avantatjoses tenen més possibilitats de sobreviure, reproduir-se i transmetre les mutacions a la seva descendència.
La selecció natural actua per eliminar les mutacions desavantatjoses; per tant, està actuant contínuament per protegir l’espècie de la decadència mutacional. No obstant això, la mutació desavantatjosa sorgeix a la mateixa velocitat a la qual la selecció natural l’elimina, de manera que les poblacions mai estan completament netes de formes mutants desavantatjoses dels gens. Aquestes mutacions que no resulten avantatjoses poden ser l’origen de malalties genètiques que es poden transmetre a la següent generació.
[11] Veure el complet article sobre Lamarck a la Wikipèdia francesa, http://fr.wikipedia.org/wiki/Jean-Baptiste_de_Lamarck
[12] Veure l’excel·lent article de la Wikipèdia a http://ca.wikipedia.org/wiki/Charles_Robert_Darwin
[13] Serge Ciccotti (2011).Tout ce que vous devez savoir pour mieux comprendre vos semblables. París. Dunod. Pàg. 356
[14] Un cas molt cèlebre és el de la falena del bedoll a Anglaterra: en aquesta espècie de papallones, la proporció d’individus clars hauria disminuït a causa de la contaminació que destruïa els líquens( blancs) que creixen en els troncs dels arbres. Efectivament, els individus clars (inicialment majoritaris), en esdevenir més visibles que els altres damunt dels troncs ennegrits, eren més fàcils per als depredadors. Les falenes fosques que ja existien abans de la contaminació esdevingueren, llavors, majoritàries. Posteriorment, el fenomen es va invertir atesa la desaparició de les indústries contaminants, cosa que va permetre el retorn del desenvolupament del liquen sobre els troncs. Aquesta història és, tanmateix, discutible ja que les falenes no es posen sobre els troncs sinó sobre les fulles dels bedolls. Les modificacions de les freqüències de les arnes fosques i clares podrien ser respostes a la toxicitat dels residus industrials abans que a la modificació del color de l’escorça dels troncs.
[15] Todd Bersaglieri, Pardis C. Sabeti, Nick Patterson, Trisha Vanderploeg, Steve F. Schaffner, Jared A. Drake, Matthew Rhodes, David E. Reich, and Joel N. Hirschhorn: «Genetic Signatures of Strong Recent Positive Selection at the Lactase Gene». The American Journal of Human Genetics, volume 74 (2004), pàgs. 1111–1120. En línia a http://www.ncbi.nlm.nih.gov/pubmed/15114531
[16] Sarah A. Tishkoff, Floyd A. Reed, Alessia Ranciaro, Benjamin F. Voight, Courtney C. Babbitt, Jesse S. Silverman, Kweli Powell, Holly M. Mortensen, Jibril B. Hirbo, Maha Osman, Muntaser Ibrahim, Sabah A. Omar, Godfrey Lema, Thomas B. Nyambo, Jilur Ghori, Suzannah Bumpstead, Jonathan K. Pritchard, Gregory A. Wray & Panos Deloukas: « Convergent adaptation of human lactase persistence in Africa and Europe». Nature Genetics, volume 39 (January 2007), pàgs. 31-40. En línea a http://www.bioquest.org/summer2007/projectfiles/lactase_persistance_Nature_gen.pdf
[17] L’Ortogènesi, evolució ortogenètica, evolució progressiva o autogènesi, és una hipòtesi biològica segons la qual la vida té una tendència innata a evolucionar d’una manera unilineal a causa d’alguna “força directora”, ja sigui interna o externa. La hipòtesi té bases filosòfiques d’essencialisme, finalisme, i de teologia, i proposa una força intrínseca que lentament transforma les espècies.
[18] Theodosius Dobzhansky i Stephen Jay Gould (1982). Genetics and the Origin of Species. Columbia University Press. 364 pàgs.
[19] Ernst Mayr (1999). Systematics and the Origin of Species from the Viewpoint of a Zoologist.Harvard University Press. 372 pàgs..
[20] George Gaylord Simpson (1984).Tempo and Mode in Evolution. Columbia University Press. 237 pàgs.
[21] Julian Huxley (1975). Evolution: The Modern Synthesis. Macmillan Pub Co. 3ª ed. 705 pàgs.
[22] Motoo Kimura (1985). The Neutral Theory of Molecular Evolution. Cambridge University Press. 384 pàgs.
[23] Dawkins, Richard (2000). El gen egoista. Barcelona. Salvat ed. 408 pàgs.
Joan Campàs Aura digital
Curs: Orígens de l’Art i evolució humana: l’homo significans
Estudis d’Arts i Humanitats de la UOC
Materials del curs en format .pdf
1. Una visió de conjunt de l’art paleolític
2. El paleolític: cronologia i referències etnoarqueològiques
3. Evolució tecnològica en el Paleolític. Referències etnoarqueològiques de l’hàbitat
4. La construcció del coneixement de l’art del Paleolític
5. El paradigma d’Henri Breuil i la seva crisi
6. L’estructuralisme de Leroi-Gourhan i Annette Laming-Emperaire
7. Del xamanisme a l’art com expressió de l’organització social
8. El procés evolutiu: Darwin i la selecció natural
9. Darwin i la selecció sexual. El cicle menstrual
10. La selecció sexual: la competència espermàtica
11. La selecció sexual: l’orgasme femení
12. Sobre els orígens de l’art: les coalicions de dones pintades
13. Una perspectiva ecològica de l’evolució humana
14. Els primers hominoides
15. Els homínids: els preaustralopitecins
16. Els hominins: els australopitecins
17. Paranthropus
18. Els primers Homo: Homo habilis
19. El poblament d’Euràsia: l’homo ergaster/erectus
20. Homo antecessor, rhodesiensis, heidelbergensis i floresiensis
21. L’Homo neanderthalensis
22. L’Homo sapiens
23. Sobre el concepte d’art
24. Les primeres expressions artístiques
25. Sobre els orígens de l’art del Plistocè
26. L’art rupestre del Plistocè
27. Temes i distribució de l’art parietal
28. L’art moble del Plistocè
29. Temes de l’art mobiliari
30. Art mobiliari del Paleolític Superior. Interpretacions
31. Sobre els orígens del llenguatge